Molar Development and Crown Areas in Early Australopithecus
|
|
|
- Annice Dean
- 6 years ago
- Views:
Transcription
1
2 AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 000: (2012) Molar Development and Crown Areas in Early Australopithecus Rodrigo S. Lacruz, 1 * Fernando V. Ramirez Rozzi, 2 Bernard A. Wood, 3,4 and Timothy G. Bromage 5,6 1 Center for Craniofacial Molecular Biology, Herman Ostrow School of Dentistry, University of Southern California, Los Angeles, CA UPR 2147 CNRS, 44 rue de l Amiral Mouchez, Paris 75014, France 3 Center for the Advanced Study of Hominid Paleobiology, George Washington University, Washington, DC Department of Anthropology, George Washington University, Washington, DC Department of Biomaterials and Biomimetics, New York University College of Dentistry, New York, NY Department of Basic Science and Craniofacial Biology, New York University College of Dentistry, New York, NY KEY WORDS Australopithecus; molar crown area; crown formation; Plio-Pleistocene ABSTRACT Recent studies suggest that the hypodigms representing the two earliest Australopithecus (Au. anamensis and Au. afarensis) form an ancestor-descendant lineage. Understanding the details of this possible transition is important comparative evidence for assessing the likelihood of other examples of ancestor-descendant lineages within the hominin clade. To this end we have analyzed crown and cusp base areas of high resolution replicas of the mandibular molars of Au. anamensis (Allia Bay and Kanapoi sites) and those of Au. afarensis (Hadar, Laetoli, and Maka). We found no statistically significant differences in crown areas between these hypodigms although the mean of M 1 crowns was smaller in Au. anamensis, being the smallest of any Australopithecus species sampled to date. Intraspecies comparison of the areas of mesial cusps for each molar type using Wilcoxon signed rank test showed no differences for Au. anamensis. Significant differences were found between the protoconid and metaconid of Au. afarensis M 2 s and M 3 s. Furthermore, the area formed by the posterior cusps as a whole relative to the anterior cusps showed significant differences in Au. afarensis M 1 s and in Au. anamensis M 2 s but no differences were noted for M 3 s of either taxon. Developmental information derived from microstructural details in enamel shows that M 1 crown formation in Au. anamensis is similar to Pan and shorter than in H. sapiens. Taken together, these data suggests that the overall trend in the Au. anamensis-au. afarensis transition may have involved a moderate increase in M 1 crown areas with relative expansion of distal cusps. Am J Phys Anthropol 000: , VC 2012 Wiley Periodicals, Inc. The two earliest Australopithecus species, Au. anamensis and Au. afarensis, are widely regarded as examples of anagenetic change within a single evolving lineage (Kimbel et al., 2006; Leakey et al., 1995; Ward et al., 2001, 2010; White et al., 2006; Haile-Selassie, 2010; Haile-Selassie et al., 2010). Indeed, recent reports of Australopithecus from Woranso-Mille, Ethiopia suggest that any distinctions between the Au. anamensis and Au. afarensis hypodigms are so trivial that there are few morphological grounds for their being separate taxa (Haile-Selassie, 2010; Haile-Selassie et al., 2010), but if the present specific distinction is maintained then the Woranso-Mille fossils would be best ascribed to Au. anamensis (Haile-Selassie, 2010). Comparisons of dental characters between these hypodigms have not yet included details of molar crown and cusp base areas and only limited information is available concerning molar development (Wood et al., 1983; Beynon and Wood, 1987; Suwa et al., 1994; Dean et al., 2001; Lacruz et al., 2008; Wood, 2010). The aims of this study are to: (1) present a comparative analysis of measured crown and cusp base areas in the molars of Au. anamensis and Au. Afarensis, and (2) present data on molar development based on exposed microstructural features. These data are used to analyze aspects of molar evolution between these two hypodigms. THE EARLIEST AUSTRALOPITHECUS The chronologically oldest species of the genus Australopithecus is Au. anamensis (Leakey et al., 1995; Ward et al., 2001) (but see below). Until recently, evidence of the dentition of Au. anamensis was known from sediments dated to 4.2 to 3.9 Ma at Kanapoi, Allia Bay, and Asa Issie (Leakey et al., 1995; White et al., 2006). The hypodigm of the younger taxon Au. afarensis is largely made up of material recovered from Hadar, Laetoli, and Maka (White, 1977; Johanson et al., 1982; White et al., 2000), plus specimens recovered more recently from Dikika (Alemseged et al., 2005). Fragmentary remains have been recovered from a number of Grant sponsor: The Leakey Foundation. *Correspondence to: Rodrigo Lacruz, University of Southern California, Center for Craniofacial Molecular Biology, 2250 Alcazar St., CSA 103, Los Angeles, CA 90033, USA. rodrigo@usc.edu Received 29 November 2011; accepted 9 April 2012 DOI /ajpa Published online in Wiley Online Library (wileyonlinelibrary.com). VC 2012 WILEY PERIODICALS, INC.
3 2 R.S. LACRUZ ET AL. other Kenyan and Ethiopian sites, and that taxon may also have been found in Chad (e.g. Kimbel and Delezene, 2009). The temporal span of the hypodigm is between about 3.7 and 3.0 Ma (Kimbel and Delezene, 2009). There has been strong support for the proposal that Au. anamensis and Au. afarensis are related by anagenesis as an ancestral-descendant lineage (Leakey et al., 1995; Ward et al., 2001; White et al., 2006, 2009; Haile-Selassie, 2010; Haile-Selassie et al. 2010). This hypothesis has been supported by the results of numerical cladistic analysis (Strait and Grine, 2004), and analysis of dentognathic characters (Kimbel et al., 2006). More recently, this hypothesis has been bolstered by the discovery of hominin fossils at Woranso-Mille in Ethiopia that date to between 3.57 and 3.8 Ma i.e., to the temporal interval between the hypodigms of Au. anamensis and Au. afarensis (Haile-Selassie, 2010; Haile-Selassie et al., 2010). These remains preserve a suite of morphological characters some of which are intermediate between Au. anamensis and Au. afarensis and some of which are shared by one or both of those taxa (Haile-Selassie, 2010; Haile- Selassie et al., 2010). Based on the Woranso-Mille evidence, distinctions between Au. anamensis and Au. afarensis were deemed confusing and unwarranted (Haile- Selassie, 2010). However, for the purpose of this study, we have maintained the taxonomic separation of these two hypodigms. Crown areas and enamel development The overall crown areas and the areas of the individual cusp boundaries of mandibular molars have been used to assess patterns of morphological variation within Plio-Pleistocene hominin species (e.g., Wood et al., 1983; Suwa et al., 1994). The methods used in these studies have been adapted for the analysis of the crown morphology of premolars (Wood et. al., 1983; Suwa, 1988) and maxillary molars (Bailey, 2004; Moggi-Cecchi and Boccone, 2007; Grine et al., 2009; Quam et al., 2009). In addition to the crown base area data, this study also includes evidence about enamel microstructure. The enamel microstructure of primates and that of many other mammalian taxa preserves daily (or short period) growth markings called cross-striations, which represent a day s worth of matrix secretion by the enamel forming cells, or ameloblasts. Besides cross-striations, enamel preserves approximately circaseptan long period growth markings called striae of Retzius, which may form perikymata where they reach the lateral enamel surface (Boyde, 1989). The number of cross striations between adjacent striae is known as the periodicity, or as the striae repeat interval and the distance between the cross-striations corresponds to the amount of enamel added to the thickness of the crown in a day, also known as the daily secretion rate. Further details on these markings can be found in Boyde (1989). MATERIALS Data were collected from the originals of the teeth of Au. anamensis reported in Leakey et al. (1995) and Ward et al. (2001). The Au. anamensis samples from Asa Issie and Woranso-Mille were not included in this analysis. A total of 17 molars of Au. anamensis teeth were analyzed (Appendix). The sample consists of M1 (n 5 7); M2 (n 5 6) and M3 (n 5 4). Because our cast collection of Au. afarensis molars was incomplete, we obtained a more complete data set kindly provided by Dr. Gen Suwa (referred to hereafter as the GS data set) which was originally reported in Suwa et al., (1994) (Appendix). In addition to these data, the Maka sample (White et al., 2000) was included, and for consistency, we used values for the Maka sample measured by GS. A total of 48 teeth of Au. afarensis were analyzed, which excludes the Dikika sample that was not examined for this study. METHODS There are subtle differences in the methods used to measure cusp and cusp boundaries in molars (Wood et al., 1983; Suwa et al., 1994; Bailey et al., 2004). The main differences are in the orientation or placing of the specimen so as to maximize the occlusal crown area. Whereas Wood et al. (1983) used the cervical line as a plane of reference; others have used the occlusal fovea to maximize the occlusal crown area (Suwa et al., 1994, 1996). Despite these differences in the method, studies suggest that it has no significant impact on the resulting measurements (Suwa et al., 1994; Bailey et al., 2004; Grine et al., 2009). In our study we followed the protocol described in Suwa et al., (1994). For Au. afarensis we used the GS data set on cusp and crown areas. For Au. anamensis, cusp areas were measured from images taken on high resolution casts of the specimens made by us using Coltène President in its light and putty variant. Casts were photographed using a Zeiss stereo microscope coupled with Nikon Coolpix 4500 at 63 magnification. Total crown base areas (CBA) as well as individual cusp areas were measured using ImageJ software. Individual cusp base areas were measured following Wood et al. (1983) in which surface areas of accessory cusps were divided equally and added to the adjacent principal cusps. Crowns in which wear had removed substantial lengths of the primary or secondary fissures or specimens that required extensive reconstruction of the crown outline were not included in this analysis. Each specimen was measured twice and the mean of the two measurements was taken to be the value for that specimen. If antimeres were available, the mean of both areas was used. To assess potential differences between Au. anamensis and Au. afarensis molars using the GS data set for Au. afarensis, we analyzed interobserver error differences by comparing the same Au. afarensis molars measured by us with measurements of the same specimens made by GS. Two tailed Student s t-test was used to assess these differences. Univariate analysis of cusp and crown areas includes basic descriptive statistics as well as the nonparametric Mann-Whitney U test for comparing the cusp and crown base areas between Au. anamensis and Au. afarensis. We compared not only the total crown base area (CBA) of the molars but also the absolute sizes of the individual cusps and their relative contribution to the composition of the occlusal surface of the whole crown. To compare differences in cusp size within the molar series of each taxon (i.e., to compare the protoconid and metaconid of each tooth type in each species), or to compare the combined areas of the distal cusps relative to anterior cusps, we employed the nonparametric Wilcoxon signed rank test. Molar development Materials. Two naturally fractured Au. anamensis molars, the M 1 KNM-KP 31712j and a possible M 1

. Methods. We have previously described in detail the protocol for estimating crown formation time in hominin molars (Lacruz et al.")
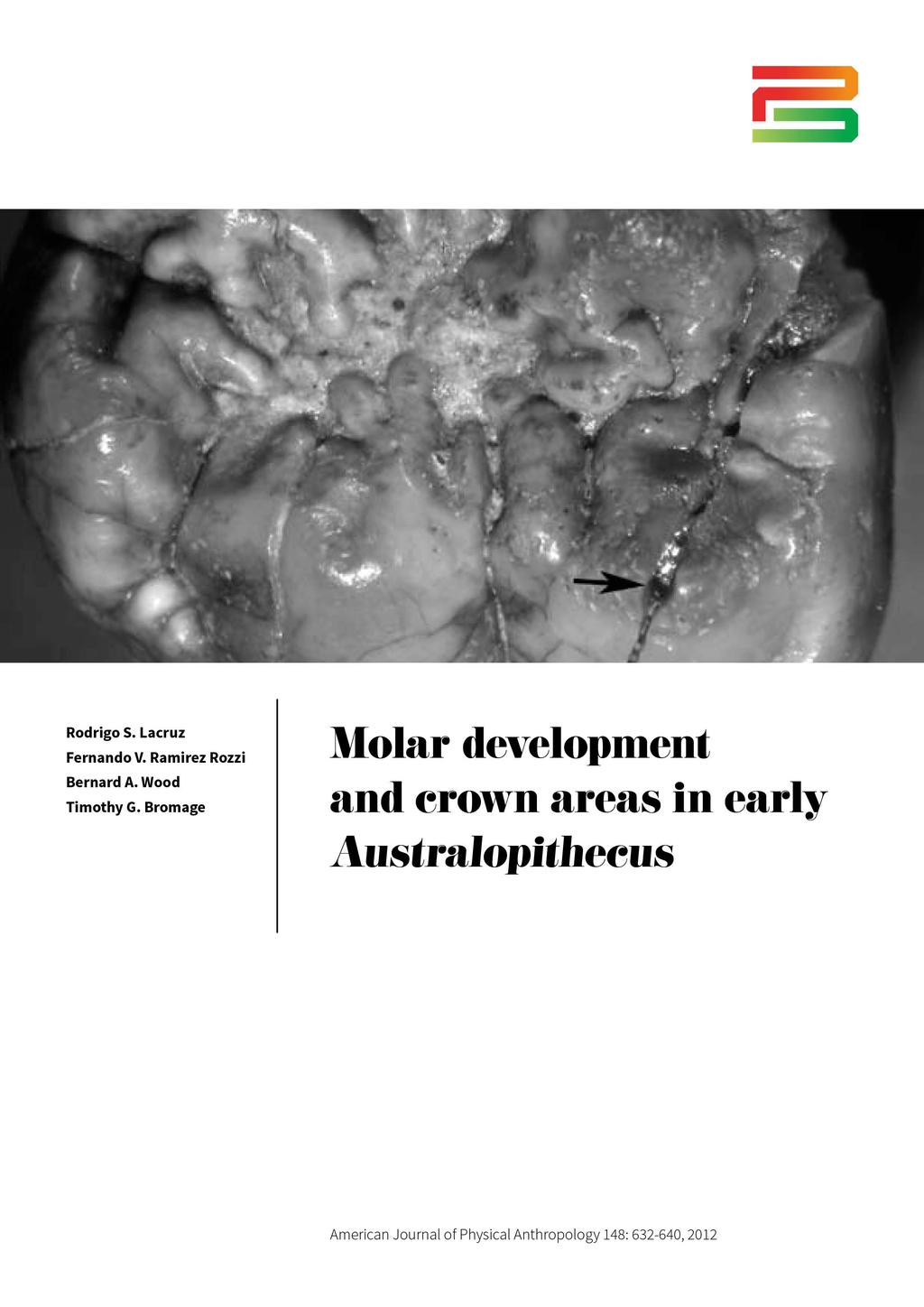
4 MOLAR DEVELOPMENT IN EARLY AUSTRALOPITHECUS 3 Fig. 1. A: The occlusal view of KNM-ER 30749, an incomplete left M 1 (?) crown showing a fracture on the metaconid cusp tip (arrowed). B: Lateral view of the metaconid indicating by a white line the approximate area where we measured the cuspal enamel thickness. Note the concave shape of the Hunter Schreger bands (HSB) in (B) in this specimen from the younger Au. anamensis site of Allia Bay, which contrasts with the shape of HSB described in some Au. afarensis molars (Lacruz and Ramirez Rozzi, 2010). Methods. We have previously described in detail the protocol for estimating crown formation time in hominin molars (Lacruz et al., 2006; Lacruz and Ramirez Rozzi, 2010). Following the method of Reid et al., (1998) the lateral perikymata or striae of Retzius are counted and the total number is multiplied by the periodicity, which provides the lateral formation time of the crown. For the occlusal or cuspal enamel we used the method originally described in Beynon et al. (1991). The linear enamel thickness is measured from a point just below the gnarled enamel at the EDJ and the region in the outer enamel where the last lateral stria is identified. This measurement is then divided by the average value of the cross striation length, providing the cuspal or occlusal formation time. The average values of cuspal cross striations for Au. anamensis and Au. afarensis were reported in Lacruz et al. (2008). Crown formation time is the sum of lateral and cuspal enamel. F1 Fig. 2. Occlusal view of KNM-KP 31712j, an M 1 of Au. anamensis. The protoconid cusp (arrowed) is perfectly fractured naturally almost at the tip as seen in the image. We were able to unglue this specimen to study its microstructure. (KNM-ER 30749; Ward et al., 2001) were used to estimate cusp and crown formation time. These specimens were selected because they showed natural fractures that passed near the dentine horn broadly oriented in the buccolingual plane (Figs. 1A,B and 2). For detailed descriptions of each specimen see Ward et al., (2001; p. 293 [KNM-KP 31712j] and p. 325 [KNM-ER 30749]). This latter specimen had been previously sectioned and cusp formation time for the hypoconid reported in Ward et al. (2001). Here we report on the metaconid cusp formation of this molar and on the protoconid of KNM- KP31712j. RESULTS Interobserver error Table 1 shows descriptive statistics for the Au. afarensis molars measured by GS and by us comparing both data sets. Two-tailed Student s t-test reveals no significant differences between measurements of the same samples made by different observers using a similar method. In fact, our results and those obtained by GS are strikingly similar when the overall mean values are considered. However, differences in individual cusp measurements differed by as much as 25% in some cases, although most commonly the differences were around 10 15%. Crown and cusp base areas Table 2 shows descriptive statistics of individual cusps and crown base areas of lower molars of Au. anamensis
5 4 R.S. LACRUZ ET AL. TABLE 1. Interobserver error measurements were assessed using t-test comparison of the means of the cusp and crown areas for Au. afarensis molars measured by us and those measured in Suwa et al. (1994) Protoconid Metaconid Hypoconid Entoconid Hypoconulid Total area M1 this study 34.3 (5.8) n (5.9) n (3.7) n (4.1) n (4.1) n (15.8) n 5 8 M1 GS 32.4 (3.7) n (4.47) n (2.5) n (5.5) n (2.8) n (14.7) n 5 8 P value P ; n 5 6 P ; n 5 6 P ; n 5 6 P ; n 5 6 P ; n 5 6 P ; n 5 8 M2 this study (7.4) n (8.2) n (5.7) n (2.9) n (6.1) n (20.2) n 5 8 M2 GS (7.1) n (7.9) n (4.0) n (1.8) n (6.5) n (19.85) n 5 8 P value P ; n 5 6 P ; n 5 6 P ; n 5 6 P ; n 5 6 P ; n 5 6 P ; n 5 8 M3 this study (4.0) n (2.4) n (4.3) n (5.8) n (6.6) n (16.1) n 5 7 M3 GS (6.5) n (4.3) n (3.6) n (5.1) n (7.2) n (17.1) n 5 7 P value P ; n 5 5 P ; n 5 5 P ; n 5 5 P ; n 5 5 P ; n 5 5 P ; n 5 7 and Au. afarensis. Au. anamensis M 1 crown base areas are somewhat smaller than those of Au. afarensis showing smaller individual cusp means, which in the case of entoconid and hypoconulid, were significantly different (P \ 0.05). Averages for M 2 s crowns were slightly higher in Au. anamensis whereas M 3 s were moderately smaller. Significant differences were observed for M 2 hypoconulids and for M 3 entoconids (Table 2). A graphical representation (box plots) of the total crown base areas of molars is shown in Figure 3. The Wilcoxon signed rank test was used to compare the absolute size of the principal mesial cusps of each molar type between the hypodigms (Table 3). For Au. anamensis, no statistical differences between the protoconid and metaconid were identified for any of the molar types. For Au. afarensis, no differences were identified in M 1 s, whereas M 2 s and M 3 s showed statistical differences between the main cusps. Despite absolute differences between the areas of the protoconid and metaconid in Au. afarensis M 2 s and M 3 s, Table 4 shows that there were no major changes in the contribution of individual cusp base areas to the total CBA between the two Australopithecus hypodigms. Table 3 also shows results of Wilcoxon signed rank test comparing the area formed by the combined measurements of the posterior cusps relative to the areas formed by the anterior cusps. For M 1 s, the increase in the areas formed by the posterior cusps was significant in Au. afarensis, but for M 2 s, an increase in the areas formed by the posterior cusps was significant in Au. anamensis. No differences were identified in M 3 s in either hypodigm. Molar development Using the method described in Lacruz et al. (2008) we were able to obtain a periodicity of 7 days for the Au. anamensis specimen KNM-KP 31712j, which together with the periodicities for two other Au. anamensis reported in Ward et al. (2001), brings to a total of three the number of Au. anamensis teeth with a known periodicity; in all three cases it is 7 days. Thus, we use the 7 day periodicity to estimate the cuspal and crown formation times of Au. anamensis M 1 s (Table 5); the former were calculated for the metaconid (KNM-ER 30749) and protoconid (KNM-KP 31712j). We used the average of the daily secretion rates from Lacruz et al. (2008) (i.e., a mean occlusal DSR of 4.7 lm). KNM-ER was slightly worn and we estimated 200 lm of missing cuspal enamel should be added to our estimations of linear enamel thickness. KNM-KP 31712j is described as an unworn M 1 crown (Ward et al., 2001, p. 293), but in this case we added 100 lm as the fracture plane appears to have missed the very tip of the cusp (Fig. 2) (Table 5). Cusp formation times for each cusp are 2.2 years for the metaconid of KNM-ER and 2.1 years for the protoconid of KNM-KP 31712j. DISCUSSION Prior to this study, only qualitative descriptions of cusp areas of Au. anamensis were available (Ward et al., 2001). For Au. afarensis, cusp and crown base areas were quantified and reported in Suwa et al., (1994) and Bromage et al., (1995). Our analysis of Au. afarensis molars presented here differs from previous studies in that we have included the Maka dental sample that was reported after the publication of Suwa et al. s and Bromage et al. s analyses. Interobserver differences between our analysis and that reported by Suwa et al., (1994) using the same specimens reveal very minor differences in results. Although minor differences were noted between the means largely ranging from 10 to 15%, no statistically significant differences were identified (Table 1). This suggests that we can confidently use the GS data on Au. afarensis to compare it with our data on Au. anamensis molars. Differences in crown base areas between Au. anamensis and Au. afarensis largely concern the smaller M 1 sof Au. anamensis (P ) (Table 2). One of the Au. anamensis M 1 specimens (KNM-KP 30500) was markedly larger than the remaining M 1 of this group available for study and was larger than all the known Au. afarensis M 1 s. It is also noticeable that the crown of the M 2 of KNM-KP is also larger than all of the Au. afarensis M 2 s. Box plots of crown areas are shown in Figure 3. Comparing the average values of the Au. anamensis crown base areas with the mean values for Au. africanus in Suwa et al., (1994) and Bromage et al., (1995), Au. anamensis crown areas are smaller than the Au. africanus mean for each molar type (Fig. 4). Compared with the East and southern African Paranthropus samples, Au. anamensis crown base area values are smaller for all tooth types. When compared with the mean of the values of the early Homo group reported by Suwa et al., (1994), Au. anamensis has moderately larger M 2 and similar M 3 mean crown base areas, whereas the M 1 s are smaller in the Au. anamensis hypodigm. The picture that emerges from these comparisons is that the Au. anamensis hypodigm has the smallest M 1 crown areas among the early Australopithecus samples analyzed to date, a finding consistent with metrical data on standard MD-BL measurements (Suwa et al., 2009; Kimbel and Delezene, 2009). Although some variation on a specimen by specimen basis is noted, Figure 4 also indicates that the overall trend in crown base area size is M 1 \ M 2 [
6 MOLAR DEVELOPMENT IN EARLY AUSTRALOPITHECUS 5 TABLE 2. Mean, range, and standard deviation for cusp and total crown areas of lower molars of Au. anamensis and Au. afarensis Protoconid Metaconid Hypoconid Entoconid Hypoconulid Total crown ( ) (24.5) N ( ) (3.8) 19.1 ( ) (2.4) 23.5 ( ) (5.9) 27.2 ( ) (3.6) Au. anamensis M ( ) (5.8) ( ) (15.4) N ( ) (2.8) N ( ) (4.9) N ( ) (2.4) N ( ) (4.4) N 5 7 Au. afarensis M ( ) (3.6) N 5 7 P values P P P P P P * ( ) (18.7) N ( ) (1.4) 25.0 ( ) (4.2) 34.4 ( ) (5.2) 39.8 ( ) (4.3) Au. anamensis M ( ) (7.6) ( ) (20.9) N ( ) (4.9) N ( ) (4.0) N ( ) (4.4) N ( ) (7.5) N 5 10 Au. afarensis M ( ) (7.4) N 5 10 P values P P P P P P ( ) (12.8) N ( ) (7.7) N ( ) (2.9) N ( ) (4.2) N ( ) (2.1) N 5 3 Au. anamensis M ( ) (1.9) N ( ) (15.9) N ( ) (5.4) N ( ) (4.1) N ( ) (4.1) N ( ) (5.4) N 5 10 Au. afarensis M ( ) (6.5) N 5 10 P values P P P P P P P values indicate results of Mann-Whitney U test comparing values of each cusp between hypodigms. M 3 in Au. anamensis and Au. africanus. This is a similar trend to that described for early Homo (Suwa et al., 1994). For the Au. afarensis hypodigm, M 2 crown base area is moderately smaller than M 3 (Fig. 4), as was also observed in P. boisei (Wood et al., 1983; Suwa et al., 1994). Molar crown development Information on the timing of the development of Au. anamensis molars was provided in Ward et al. (2001). The cusp formation times published in Ward et al. (2001) are as follows: KNM-KP protocone years and KNM-ER hypoconid years. Ward et al. (2001) indicated that "the greater time for cusp formation in the KNM-ER hypoconid than the KNM-KP protocone suggests that this specimen belongs more distal in the tooth row than does KNM-KP " Indeed M 1 s have shorter crown formation than M 2 s and M 3 s in humans and in chimpanzees (Reid and Dean, 2006; Smith et al., 2007). However, in Ward et al., (2001; p. 325) it is indicated that KNM-ER is a left M 1. Given the uncertainty concerning KNM-ER and because of a method recently reported to assess crown formation time based on cusp formation time (see below), it is perhaps more informative to focus our attention on the protoconid of KNM-KP 31712j (M 1 ) to assess crown formation. The cusp formation time obtained was 2.1 years, which is in line with the values reported by Ward et al., (2001) for a maxillary protocone. We have previously estimated molar crown formation times in molars of Au. afarensis, Au. Africanus, and P. robustus from cusp formation times using a method originally described by Ramirez Rozzi (1993) in an analysis of Ethiopian specimens from the Omo Formation (Lacruz et al., 2006; Lacruz and Ramirez Rozzi, 2010). In this study we use a different method. The reason to do so is the difficulty in following the last cervical stria to its corresponding perikyma and following this to the distal moiety of the molar, as we had previously done (Ramirez Rozzi, 1993; Lacruz et al., 2006). Thus we relied on the results of a recent study of modern human M 1 crown development that showed that the time taken to develop the protoconid approximates (with a small, c. 7%, margin of error) to the time taken to form the whole crown (Mahoney, 2008). Using this relationship and the estimate of the time to form the protoconid of KNM-KP 31712j (2.1 years or 759 days), we can obtain an estimate of 2.2 years (759 days 1 7% or 53 days days) for the formation time of the entire crown. We did not use the same method to assess crown formation time in KNM-ER because inferring total crown formation time based on metaconid formation times had a noticeably greater error (Mahoney, 2008). However, the crown formation time for this molar based on our results and those reported in Ward et al. (2001) can be estimated to be between 2.2 and 2.7 years. The estimated crown formation time of 2.2 years for KNM-KP 31712j (M 1 ) is similar to the reported average value of chimpanzee (Pan troglodytes) M 1 crown formation (Smith et al., 2007) suggesting that this may be the symplesiomorpic condition for early hominins. The crown formation time is also shorter than the corresponding values for Homo sapiens (Reid and Dean, 2006). A cautionary note should be included here. We had originally reported on the meta-
7 6 R.S. LACRUZ ET AL. Fig. 3. Box plots of total crown areas for each molar type in lower molars of Au. anamensis and Au. afarensis. conid cusp formation time of A.L which was described by Johanson et al. (1982) as an M 1. However, it has come to our attention that this tooth may be best classified as an M 2 and that the cusp analyzed by us was not a metaconid but an entoconid (Gen Suwa pers. comm.). Therefore, data obtained here for Au. anamensis M 1 s may not be directly compared with like-tooth type in Au. afarensis. It is well known that the timing of M 1 eruption is highly correlated with life history-related variables (i.e., cranial capacity) (Smith, 1991). For hominins, however, data on molar eruption times are scarce (e.g., Dean, 2010) but in the absence of such information, M 1 crown formation time could possibly be used as a broad indicator of hominin life history, although this issue remains a subject of study (Macho, 2001; Kelley and Smith, 2003; Schwartz et al., 2005). In hominins, this approach is TABLE 3. Top: Intraspecies comparison of the areas of the main anterior cusps (protoconid vs. metaconid) for each molar type in Au. anamensis and Au. afarensis lower molars using Wilcoxon signed rank test; Bottom: Comparison of the relative areas of the individual posterior cusps combined with the combined areas of the anterior cusps for each molar type using Wilcoxon signed rank test Protoconid vs. metaconid Hypodigm M1 M2 M3 Au. anamensis P P P Au. afarensis P P P Anterior vs. posterior cusps M1 M2 M3 Au. anamensis P P P Au. afarensis P P P hampered because of the few data that are available on M 1 crown formation time (Lacruz and Ramirez Rozzi, 2010). Despite these uncertainties, M 1 crown formation time can be used to predict the age at death of certain individuals (e.g. Lacruz et al., 2005). Using our reported M 1 crown formation for Au. anamensis (2.2 years, Table 5), together with the average rate of root growth in Au. anamensis, apes or in modern humans of between 1 and 2 mm of the first root formed (c. 6.0 microns/day) (Dean, 2010), this information may be used to estimate age at death of juvenile Au. anamensis fossils in the future. Furthermore, because the time invested by each species in forming a tooth depends on the schedule of growth and development of that species and the time available to do this (e.g. Bromage, 1987; Dean, 2006; Lacruz et al., 2008) the data on crown formation time presented here and the reported data on Au. anamensis root formation (Dean, 2010) suggests potentially similar developmental schedules for Au. anamensis and Pan. This matches what has been proposed some time ago for Au. afarensis (Bromage and Dean, 1985; Anemone, 2002). Morphological changes in Australopithecus molars Among the differences identified between the Au. anamensis and Au. afarensis fossils, including the Woranso- Mille material, the mandibular molars of Au. anamensis are described as being lower crowned than those of Au. afarensis and possessing "sloping buccal sides" (Leakey et al., 1995). The maxillary molars possess "trigons much wider than talons" (Leakey et al., 1995). However, the size of the permanent postcanine teeth was considered to be similar in the two species (Haile-Selassie et al., 2010; Ward et al., 2001). The occlusal morphology of mandibular molars (i.e., as judged by the observed size of main cusps) was also described as being similar in these taxa although no quantitative data was reported (Ward et al., 2001). Descriptions of the occlusal areas of lower molars indicated that the largest cusps in Au. anamensis were the protoconid and metaconid (Ward et al., 2001, p.350). It was also noted that the fissure patterns of the Au. anamensis molars exhibited enough variation to suggest caution about the use of such features for taxonomic purposes (Ward et al., 2001, p. 350). The results of the quantitative analyses presented here, which do not include the Woranso-Mille or Asa Issie Au. anamensis, show that the crown base area values of Au. anamensis M 1 s are smaller than those for Au. afarensis, whereas M 2 are slightly bigger and have similar M 3 crown. Few statistically significant differences were identified for any of the crowns of the molar types although differences were identified in some of the distal cusps (entoconid and hypoconulid) of all molar positions. These differences may be related to the complexities of the distal moiety of lower molars which makes difficult to clearly identify the boundaries of the distal cusps TABLE 4. Percentage that each individual cusp area contributes to the total crown area in lower molars of Au. anamensis and Au. afarensis Hypodigm Molar Protoconid Metaconid Hypoconid Entoconid Hypoconulid Au. anamensis M1 23.6% (sd 2.3) 23.3% (sd 1.9) 20.6% (sd 2.1) 16.7% (sd 3.0) 15.7% (sd 2.4) Au. afarensis M % (sd 1.4) 22.0% (sd 0.9) 21.5% (sd 0.9) 17.4% (sd 1.9) 15.7% (sd 1.1) Au. anamensis M2 24.9% (sd 2.7) 23.1% (sd 0.9) 19.5% (sd 1.1) 15.6% (sd 1.5) 16.7% (sd 1.6) Au. afarensis M2 26.1% (sd 1.9) 22.9% (sd.18) 19.7% (sd 1.5) 16.1% (sd 2.1) 15.2% (sd 1.9) Au. anamensis M3 26.2% (sd 2.4) 22.5% (sd 2.0) 16.4% (sd 2.0) 13.4% (sd 4.1) 21.4% (sd 3.8) Au. afarensis M3 26.1% (sd 1.6) 22.8% (sd 2.7) 17.0% (sd 1.7) 17.1% (sd 1.6) 17.0% (sd 2.6)
8 MOLAR DEVELOPMENT IN EARLY AUSTRALOPITHECUS 7 TABLE 5. Crown formation time in Au. anamensis molars Locality Allia Bay Kanapoi Specimen KNM-ER KNM-KP 31712j Cusp Metaconid Protoconid LET 1,147 lm 1,100 lm C/LET 1,347 lm 1,200 lm Lateral/SR Periodicity 7 7 AV Oc/DSR (4.7 lm) (4.7 lm) LFT/d OFT/d Cusp FT/years Total CFT/years? 2.2 individual cusp areas, are indicative of changes in the properties of foods consumed (Lucas et al., 1986). In keeping with this notion and with previous studies (Teaford and Ungar, 2000; Ward et al., 2001, 2010; White et al., 2006), a modest increase in M 1 crown base area from Au. anamensis to Au. afarensis is consistent with a modest shift in dietary adaptations between these two groups (but see Grine et al., 2006; Ungar et al., 2010). This adaptive shift appears to be supported by additional material of Au. anamensis that is not yet fully described (Ward et al., 2010). This study contributes to a better understanding of the evolution of mandibular molars within Australopithecus and provides additional evidence about the nature of the relationships between Au. anamensis and to Au. afarensis. ACKNOWLEDGMENT Fig. 4. Comparison of the averages of crown areas in Australopithecus molars. The molars of Au. anamensis are smaller than other Australopithecus. These values are, however similar, to those reported for early Homo (Suwa et al., 1994). It is also noticeable that there has been an increase in M 1 crown areas from the oldest Australopithecus (Au. anamensis) to the youngest species of the genus (Au. africanus). Au. africanus data were obtained from Suwa et al. (1994). (Suwa et al. 1994, 1996; Ward et al., 2001; Bailey et al., 2004). Although Wilcoxon signed rank test showed no statistically significant differences between the protoconid and metaconid in the Au. anamensis molar series, we identified significant differences for Au. afarensis M 2 s and M 3 s (Table 3). Furthermore, when the contribution of the individual cusps areas is considered in relation to the whole crown for all molar types the results for the two taxa were similar (Table 4). Table 2 also indicates that for the M 2 s and M 3 s of Au. anamensis and Au. afarensis, the protoconid[metaconid[hypoconid, whereas in the M 1 s the mesial cusps are sub-equal in size. Finally, the expansion of the distal moiety of Au. afarensis M 1 s was significant relative to the area formed by the anterior cusps. For M 2 s this is noted in the Au. anamensis hypodigm only, whereas no differences were noted in M 3 s for either hypodigm. These data indicate that for the lower molars, the transition from Au. anamensis to Au. afarensis may have included a moderate increase in M 1 crown area with expansion of the distal moiety of this tooth. CONCLUSION Our analysis suggests that there may have been a moderate increase in M 1 crown base area from Au. anamensis to Au. afarensis. Previous commentators have suggested that changes in tooth crown size, which we here extend to include changes in overall crown and The authors express their gratitude to the Kenyan and Ethiopian Governments, the National Museums of Kenya and the staff at the Ethiopian National Museum, Addis Ababa, and to Mamitu Yilma and Emma Mbua for access to the fossil material in their care. They are indebted to Meave Leakey for access to the Au. anamensis material. Gen Suwa is thanked for allowing them to use his Au. afarensis data and for critically reviewing previous drafts which substantially improved this manuscript. They also thank an anonymous reviewer for helpful comments. LITERATURE CITED Anemone RL Dental development and life history in hominid evolution. In: Minugh-Purvis N, McNamara KJ, editors. Human evolution through developmental change. Baltimore: Johns Hopkins University Press. p Alemseged Z, Wynn JG, Kimbel WH, Reed D, Geraads D, Bobe R A new hominin from the Basal Member of the Hadar Formation, Dikika, Ethiopia, and its geological context. J Hum Evol 49: Bailey SE A morphometric analysis of maxillary molar crown of Middle-Late Pleistocene hominins. J Hum Evol 47: Bailey SE, Pilbrow VC, Wood BA Interobserver error involved in independent attempts to measure cusp base areas of Pan M1s. J Anat 205: Beynon AD, Dean MC, Reid DJ On thick and thin enamel in hominoids. Am J Phys Anthropol 86: Beynon AD, Wood B Patterns and rates of enamel growth on the molar teeth of early hominids. Nature 326: Boyde A Enamel. In: Berkovitz BKB, Boyde A, Frank RM, Hohling HJ, Moxham BJ, Nalbandian J, Tonge CH, editors. Teeth. Handbook of microscopic anatomy, Vol.6. Berlin: Springer. p Bromage TG The biological and chronological maturation of early hominids. J Hum Evol 16: Bromage TG, Dean MC Re-evaluation of the age at death of immature fossil Hominids. Nature 317: Bromage TG, Schrenk F, Zonnenveld FW Palaeoanthropology of the Malawi Rift: an early hominid mandible from the Chiwondo Beds, northern Malawi. J Hum Evol 28: Dean MC Tooth microstructure tracks the pace of human life-history evolution. Proc R Soc B 273: Dean MC Retrieving chronological age from dental remains of early fossil hominins to reconstruct human growth in the past. Philos Trans R Soc Lond B Biol Sci 365: Dean C, Leakey MG, Reid D, Schrenk F, Schwartz GT, Stringer C, Walker A Growth processes in teeth distinguish modern humans from Homo erectus and earlier hominins. Nature 414:
9 8 R.S. LACRUZ ET AL. Grine FE, Smith HF, Heesy CP, Smith EJ Phenetic affinities of Plio-Pleistocene Homo fossils from South Africa: molar cusp proportions. In: Grine FE, Fleagle JG, Leakey RE, editors. The first humans: origin and evolution of the genus Homo. Dordrecht: Springer. p Grine FE, Ungar PS, Teaford MF Was the Early Pliocene hominin Australopithecus anamensis a hard object feeder? S Afr J Sci 102: Haile-Selassie Y Phylogeny of early Australopithecus: new fossil evidence from the Woranso-Mille (central Afar, Ethiopia). Philos Trans R Soc Lond B Biol Sci 365: Haile-Selassie Y, Saylor BZ, Deino A, Alene M, Latimer BM New hominid fossils from Woranso-Mille (Central Afar, Ethiopia) and taxonomy of early Australopithecus. Am J Phys Anthropol 141: Johanson DC, White TD, Coppens Y Dental remains from the hadar formation. Ethiopia: collections. Am J Phys Anthropol 57: Kelley J, Smith TM Age at first molar emergence in early Miocene Afropithecus turkenensis and life history evolution in the Hominoidea. J Hum Evol 44: Kimbel WH, Delezene LK "Lucy" redux: a review of research on Australopithecus afarensis. Am J Phys Anthropol 140:2 48. Kimbel WH, Lockwood CA, Ward CV, Leakey MG, Rak Y, Johanson DC Was Australopithecus anamensis ancestral to A. afarensis? A case of anagenesis in the hominin fossil record. J Hum Evol 51: Lacruz RS, Dean MC, Ramirez Rozzi FV, Bromage TG Megadontia, patterns of enamel secretion, and striae periodicity in Plio-Pleistocene fossil hominins. J Anat 213: Lacruz RS, Ramirez Rozzi FV Molar crown development in Australopithecus afarensis. J Hum Evol 58: Lacruz RS, Ramirez Rozzi F, Bromage TG Linear enamel hypoplasia, weaning, and age at death in the Taung child. S Afr J Sci 101: Lacruz RS, Ramirez Rozzi FV, Bromage TG Variation in enamel development of South African fossil hominids. J Hum Evol 51: Leakey MG, Feibel CS, McDougall I, Walker A New four million-year-old hominid species from Kanapoi and Allia Bay. Nature 376: Lucas PW, Corlett RT, Luke DA Postcanine tooth size and diet in anthropoids. Z Morphol Anthropol 76: Macho GA Primate crown formation time and life history evolution revisited. Am J Primatol 55: Mahoney P Intraspecific variation in M1 enamel development in modern humans: implications for human evolution. J Hum Evol 55: Moggi-Cecci J, Boccone S Maxillary molar cusp morphology of South African australopithecines. In: Bailey S, Hublin J-J, editors. Dental perspectives in human evolution: state of the art research in dental paleoanthropology. Dordrecht: Springer. p Quam R, Bailey S, Wood BA Evolution of M 1 crown size and cusp proportions in the genus Homo. J Anat 214: Ramirez Rozzi F Tooth development in East African Paranthropus. J Hum Evol 24: Reid DJ, Dean MC Variation in modern human enamel formation times. J Hum Evol 50: Reid DJ, Schwartz GT, Dean C. Chandrasekera MS A histological reconstruction of dental development in the common chimpanzee, Pan troglodytes. J Hum Evol 35: Schwartz GT, Mahoney P, Godfrey LR, Cuozzo FP, Jungers WL, Randria GF Dental development in Megaladapis edwardsi (Primates, Lemuriformes): implications for understanding life history variation in subfossil lemurs. J Hum Evol 49: Smith BH Dental development and the evolution of life history in the Hominidae. Am J Phys Anthropol 86: Smith TM, Reid DJ, Dean MC, Olejniczak AK, Martin LB Molar development in the common chimpanzee (Pan troglodytes). J Hum Evol 52: Strait DS, Grine FE Inferring hominoid and early hominid phylogeny using craniodental characters: the role of fossil taxa. J Hum Evol 47: Suwa G Evolution of the robust australopithecines in the Omo succession: evidence from mandibular premolar morphology. In: Grine FE, editor. Evolutionary history of the robust australopithecines. New York: Aldine de Gruyter. p Suwa G, Wood BA, White TD Further analysis of mandibular molar crown and cusp areas in Pliocene and early Pleistocene hominids. Am J Phys Anthropol 93: Suwa G, White TD, Howell FC Mandibular postcanine dentition from the Shungura Formation, Ethiopia: crown morphology, taxonomic allocations, and Plio-Pleistocene hominid evolution. Am J Phys Anthropol 101: Suwa G, Kono RT, Simpson SW, Asfaw B, Lovejoy CO, White TD Palaeobiological implications of the Ardipithecus ramidus dentition. Science 326:94 99 (and Supporting Online Material). Teaford MF, Ungar PS Diet and the evolution of the earliest human ancestors. Proc Natl Acad Sci USA 97: Ungar PS, Scott RS, Grine FE, Teaford MF Molar microwear textures and the diets of Australopithecus anamensis and Australopithecus afarensis. Philos Trans R Soc Lond B Biol Sci 365: Ward CV, Leakey MG, Walker A Morphology of Australopithecus anamensis from Kanapoi and Allia Bay, Kenya. J Hum Evol 41: Ward CV, Plavcan JM, Manthi FK Anterior dental evolution in the Australopithecus anamensis-afarensis lineage. Philos Trans R Soc Lond B Biol Sci 365: White TD New fossil hominids from Laetolil, Tanzania. Am J Phys Anthropol 46: White TD, Suwa G, Simpson S, Asfaw B Jaws and teeth of Australopithecus afarensis from Maka, Middle Awash, Ethiopia. Am J Phys Anthropol 111: White TD, WoldeGabriel G, Asfaw B, Ambrose S, Beyene Y, Bernor RL, Boisserie JR, Currie B, Gilbert H, Haile-Selassie Y, Hart WK, Hlusko LJ, Howell FC, Kono RT, Lehmann T, Louchart A, Lovejoy CO, Renne PR, Saegusa H, Vrba ES, Wesselman H, Suwa G Asa Issie, Aramis and the origin of Australopithecus. Nature 440: White TD, Asfaw B, Beyene Y, Haile-Selassie Y, Lovejoy CO, Suwa G, WoldeGabriel G Ardipithecus ramidus and the paleobiology of early hominids. Science 326: Wood BA Reconstructing human evolution: achievements, challenges, and opportunities. Proc Nat Acad Sci USA 107: Wood BA, Abbott SA, Graham SH Analysis of the dental morphology of Plio-Pleistocene hominids. II. Mandibular molars: study of cusp areas, fissure pattern and cross sectional shape of the crown. J Anat 137:
10 MOLAR DEVELOPMENT IN EARLY AUSTRALOPITHECUS 9 APPENDIX: LIST OF SPECIMENS USED IN THIS STUDY Au. anamensis Au. afarensis Specimen Locality Specimen Locality M1 KNM-KP Kanapoi AL Hadar KNM-KP Kanapoi AL Hadar KNM-KP Kanapoi AL200 1b Hadar KNM-KP Kanapoi AL266 1 Hadar KNM-KP Kanapoi AL288 1 Hadar KNM-ER Allia Bay AL Hadar KNM-ER Allia Bay AL333w-12 Hadar AL333w-1 Hadar AL333w-60 Hadar LH.2 Laetoli LH.3 Laetoli LH. 4 Laetoli LH.16 Laetoli M2 KNM-KP Kanapoi AL Hadar KNM-KP Kanapoi AL Hadar KNM-KP Kanapoi AL188 1 Hadar KNM-KP Kanapoi AL198 1 Hadar KNM-KP Kanapoi AL Hadar KNM-ER Allia Bay AL266 1 Hadar AL277 1 Hadar AL288 1 Hadar AL333w-1 Hadar AL333w-27 Hadar AL333w-57 Hadar AL333w-59 Hadar AL333w-60 Hadar AL400 Hadar LH.4 Laetoli LH.19 Laetoli LH.23 Laetoli MAK1 2 Maka MAK1 3 Maka MAK1 12 Maka M3 KNM-KP Kanapoi AL188 1 Hadar KNM-KP Kanapoi AL198 1 Hadar KNM-KP Kanapoi AL266 1 Hadar KNM-ER Allia Bay AL288 1 Hadar AL Hadar AL Hadar AL333w-57 Hadar AL333w-59 Hadar AL333w-32/60 Hadar AL400 1 Hadar AL366 1 Hadar LH.4 Laetoli LH.15 Laetoli MAK1 2 Maka MAK1 12 Maka
Mammals Grew 1,000 Times Larger After the Demise of the Dinosaurs
 Mammals Grew 1,000 Times Larger After the Demise of the Dinosaurs The largest land mammals that ever lived, Indricotherium and Deinotherium, would have towered over the living African Elephant. Indricotherium
Mammals Grew 1,000 Times Larger After the Demise of the Dinosaurs The largest land mammals that ever lived, Indricotherium and Deinotherium, would have towered over the living African Elephant. Indricotherium
4/20/2008. Overview. Early Human Evolution. Chronology of Hominid Evolution. Overview of Species. Epochs of the Cenozoic Era
 Early Human Evolution Overview and Chronology What makes us human? Ardipithecus and early Australopithecus Robust and gracile australopithecines Oldowan tools Overview First hominins appeared late in the
Early Human Evolution Overview and Chronology What makes us human? Ardipithecus and early Australopithecus Robust and gracile australopithecines Oldowan tools Overview First hominins appeared late in the
Journal of Human Evolution
 Journal of Human Evolution 60 (2011) 711e730 Contents lists available at ScienceDirect Journal of Human Evolution journal homepage: www.elsevier.com/locate/jhevol Evolution of the mandibular third premolar
Journal of Human Evolution 60 (2011) 711e730 Contents lists available at ScienceDirect Journal of Human Evolution journal homepage: www.elsevier.com/locate/jhevol Evolution of the mandibular third premolar
The Pliocene hominin diversity conundrum: Do more fossils mean less clarity?
 SPECIAL FEATURE: PERSPECTIVE SPECIAL FEATURE: PERSPECTIVE The Pliocene hominin diversity conundrum: Do more fossils mean less clarity? Yohannes Haile-Selassie a,b,1, Stephanie M. Melillo c, and Denise
SPECIAL FEATURE: PERSPECTIVE SPECIAL FEATURE: PERSPECTIVE The Pliocene hominin diversity conundrum: Do more fossils mean less clarity? Yohannes Haile-Selassie a,b,1, Stephanie M. Melillo c, and Denise
Lecture 10-1 Early Fossil Hominids: Bipedal Anatomy & Pre- Australopithecines and Australopithecines
 Lecture 10-1 Early Fossil Hominids: Bipedal Anatomy & Pre- Australopithecines and Australopithecines Big Questions 1. What is a hominid? 2. Why did hominids evolve from an apelike primate? 3. Who were
Lecture 10-1 Early Fossil Hominids: Bipedal Anatomy & Pre- Australopithecines and Australopithecines Big Questions 1. What is a hominid? 2. Why did hominids evolve from an apelike primate? 3. Who were
Morphometric Analysis of Hominoid Lower Molars from Yuanmou of Yunnan Province, China
 PRIMATES, 42(2): 123-134, April 21 123 Morphometric Analysis of Hominoid Lower Molars from Yuanmou of Yunnan Province, China W. LIw The Chinese Academy of Sciences L. J. HLUSKO The Pennsylvania State University
PRIMATES, 42(2): 123-134, April 21 123 Morphometric Analysis of Hominoid Lower Molars from Yuanmou of Yunnan Province, China W. LIw The Chinese Academy of Sciences L. J. HLUSKO The Pennsylvania State University
Human Evolution - Skull Analysis
 Name: Date: Human Evolution - Skull Analysis Prior Knowledge Questions (Do these BEFORE using the Gizmo.) 1. Label one of the skulls below as human and the other as a chimpanzee skull. 2. What features
Name: Date: Human Evolution - Skull Analysis Prior Knowledge Questions (Do these BEFORE using the Gizmo.) 1. Label one of the skulls below as human and the other as a chimpanzee skull. 2. What features
early hominid fossils from AFRICA
 ORIGINS MATT MAHURIN (illustration); ROBERT CAMPBELL (left); ALAN WALKER; NATIONAL MUSEUMS OF KENYA (center and right) early hominid fossils from AFRICA The year was 1965. Bryan Patterson, a paleoanthropologist
ORIGINS MATT MAHURIN (illustration); ROBERT CAMPBELL (left); ALAN WALKER; NATIONAL MUSEUMS OF KENYA (center and right) early hominid fossils from AFRICA The year was 1965. Bryan Patterson, a paleoanthropologist
The Evolutionary History of the Australopiths
 Evo Edu Outreach (2010) 3:341 352 DOI 10.1007/s12052-010-0249-6 ORIGINAL SCIENTIFIC ARTICLE The Evolutionary History of the Australopiths David S. Strait Published online: 27 July 2010 # Springer Science+Business
Evo Edu Outreach (2010) 3:341 352 DOI 10.1007/s12052-010-0249-6 ORIGINAL SCIENTIFIC ARTICLE The Evolutionary History of the Australopiths David S. Strait Published online: 27 July 2010 # Springer Science+Business
NOTES: Ch 34 - Mammals & Primate / Human Evolution ( )
") NOTES: Ch 34 - Mammals & Primate / Human Evolution (34.7-34.8) Class: MAMMALIA Mammals possess unique derived characteristics: 1) Provide young with milk (mammary glands) 2) Internal fertilization; some
NOTES: Ch 34 - Mammals & Primate / Human Evolution (34.7-34.8) Class: MAMMALIA Mammals possess unique derived characteristics: 1) Provide young with milk (mammary glands) 2) Internal fertilization; some
Hominid! Evolution: On The Origin of Humans
 What is a Hominid? Hominid! Evolution: On The Origin of Humans The term hominid is also used in the more restricted sense as hominins Humans and relatives of humans closer than chimpanzees Bipedal Modern
What is a Hominid? Hominid! Evolution: On The Origin of Humans The term hominid is also used in the more restricted sense as hominins Humans and relatives of humans closer than chimpanzees Bipedal Modern
Study Guide Primates and Human Evolution. Where do you fit into the natural world? Characteristics of Primates
 Study Guide Primates and Human Evolution Describe the traits of primates.! Classify yourself taxonomically.! What traits make you human?! Describe the evolutionary trends in hominin species over the past
Study Guide Primates and Human Evolution Describe the traits of primates.! Classify yourself taxonomically.! What traits make you human?! Describe the evolutionary trends in hominin species over the past
Development Team. Physical/Biological Anthropology. Anthropology. Principal Investigator. Paper Coordinator. Content Writer.
 Paper No. : 01 Physical/ Biological Module : 15 Development Team Principal Investigator Prof. Anup Kumar Kapoor Department of, University of Delhi Paper Coordinator Prof. Subho Roy Department of,university
Paper No. : 01 Physical/ Biological Module : 15 Development Team Principal Investigator Prof. Anup Kumar Kapoor Department of, University of Delhi Paper Coordinator Prof. Subho Roy Department of,university
Internet Assignment: Early Hominids
 ANTHRO 1-L: Biological Anthropology Lab R. Mitchell, Instructor Name: Internet Assignment: Early Hominids From the late Miocene (10-5.5 mya) to the early Pliocene (5.5-4 mya), a major adaptive shift was
ANTHRO 1-L: Biological Anthropology Lab R. Mitchell, Instructor Name: Internet Assignment: Early Hominids From the late Miocene (10-5.5 mya) to the early Pliocene (5.5-4 mya), a major adaptive shift was
A n t h r o p o l o g y
 A n t h r o p o l o g y Appreciating Human Diversity Fifteenth Edition Conrad Phillip Kottak University of Michigan McGraw-Hill 2013 McGraw-Hill Companies. All Rights Reserved. C H A P T E R EARLY HOMININS
A n t h r o p o l o g y Appreciating Human Diversity Fifteenth Edition Conrad Phillip Kottak University of Michigan McGraw-Hill 2013 McGraw-Hill Companies. All Rights Reserved. C H A P T E R EARLY HOMININS
Outline. Evolution: Human Evolution. Primates reflect a treedwelling. Key Concepts:
 Evolution: Human Evolution Primates reflect a treedwelling heritage Outline 1. Key concepts 2. Characteristics of primates 3. Prosimians and anthropoids 4. The first hominids: Ardipithecus 5. The first
Evolution: Human Evolution Primates reflect a treedwelling heritage Outline 1. Key concepts 2. Characteristics of primates 3. Prosimians and anthropoids 4. The first hominids: Ardipithecus 5. The first
ARTICLE. New species from Ethiopia further expands Middle Pliocene hominin diversity
 ARTICLE doi:10.1038/nature14448 New species from Ethiopia further expands Middle Pliocene hominin diversity Yohannes Haile-Selassie 1,2, Luis Gibert 3, Stephanie M. Melillo 4, Timothy M. Ryan 5, Mulugeta
ARTICLE doi:10.1038/nature14448 New species from Ethiopia further expands Middle Pliocene hominin diversity Yohannes Haile-Selassie 1,2, Luis Gibert 3, Stephanie M. Melillo 4, Timothy M. Ryan 5, Mulugeta
1. Primate evolution provides a context for understanding human origins
 1. Primate evolution provides a context for understanding human origins Primates are monkeys, lemurs, tarsiers and apes (including us!). Compared to other mammals Most primates have hands and feet adapted
1. Primate evolution provides a context for understanding human origins Primates are monkeys, lemurs, tarsiers and apes (including us!). Compared to other mammals Most primates have hands and feet adapted
Student Exploration: Human Evolution - Skull Analysis
 Name: Date: Student Exploration: Human Evolution - Skull Analysis Prior Knowledge Questions 1. Label one of the skulls below as human and the other as a chimpanzee skull. 2. What features did you use to
Name: Date: Student Exploration: Human Evolution - Skull Analysis Prior Knowledge Questions 1. Label one of the skulls below as human and the other as a chimpanzee skull. 2. What features did you use to
Human evolution. Fascinating subject - where did we come from? History of Primates:
 Human evolution. Fascinating subject - where did we come from? History of Primates: - evolved from shrews during Cretaceous (so an older order) about 65 mya. - Some characteristics of primates: - clavicle
Human evolution. Fascinating subject - where did we come from? History of Primates: - evolved from shrews during Cretaceous (so an older order) about 65 mya. - Some characteristics of primates: - clavicle
Student Wrap-up. Topic: Investigating Hominoid Fossils: Evidence of Evolution
 Student Wrap-up Topic: Investigating Hominoid Fossils: Evidence of Evolution Benchmark: SC.912.L.15.10 Identify basic trends in hominid evolution from early ancestors six million years ago to modern humans,
Student Wrap-up Topic: Investigating Hominoid Fossils: Evidence of Evolution Benchmark: SC.912.L.15.10 Identify basic trends in hominid evolution from early ancestors six million years ago to modern humans,
Clavicle well developed (allows increase flexibility, supports arms). Five digits, front and rear. Often thumb (and big toe) opposable.
. Five digits, front and rear. Often thumb (and big toe) opposable.") Human evolution. It d be nice to spend some time with some other groups (e.g. dinosaurs), but this just isn t possible in a survey course like this. BUT, we will spend a little time on human evolution!
Human evolution. It d be nice to spend some time with some other groups (e.g. dinosaurs), but this just isn t possible in a survey course like this. BUT, we will spend a little time on human evolution!
Cenozoic Climates. Human Evolution and Adaptation
 Cenozoic Climates Human Evolution and Adaptation Life Styles of the Merely Hominid Miocene Climates Miocene Habitats The increase in climate variability would have been evident in many regions as increased
Cenozoic Climates Human Evolution and Adaptation Life Styles of the Merely Hominid Miocene Climates Miocene Habitats The increase in climate variability would have been evident in many regions as increased
Investigating Hominoid Fossils Laboratory
 Biology I Unit V: Zoology Chapter 25-28 & DOL: Vertebrates Investigating Hominoid Fossils Laboratory Name: Date: Hour: Investigating Hominoid Fossils Laboratory Pre-Lab Discussion Because hominoid fossils
Biology I Unit V: Zoology Chapter 25-28 & DOL: Vertebrates Investigating Hominoid Fossils Laboratory Name: Date: Hour: Investigating Hominoid Fossils Laboratory Pre-Lab Discussion Because hominoid fossils
Hominid Skull Comparisons
 Hominid Skull Comparisons Visit the following website: www.humanorigins.si.edu/evidence/human-family-tree Explore the interactive Human Family Tree. What can you conclude about the evolution of humans
Hominid Skull Comparisons Visit the following website: www.humanorigins.si.edu/evidence/human-family-tree Explore the interactive Human Family Tree. What can you conclude about the evolution of humans
The Human Animal. The Human Timescale. Geological Timescale. Millions of Years. Periods Jurassic. Major events
 The Human Animal The Human Timescale Geological Timescale Millions of Years Periods Permian Triassic Jurassic Cretaceous Tertiary Quat. Major events Dinosaurs Evolve and Expand Start of Age of Reptiles
The Human Animal The Human Timescale Geological Timescale Millions of Years Periods Permian Triassic Jurassic Cretaceous Tertiary Quat. Major events Dinosaurs Evolve and Expand Start of Age of Reptiles
The Human Animal. The Human Timescale. Geological Timescale. Millions of Years. Periods Permian Triassic Jurassic Cretaceous Tertiary Quat.
 The Human Animal 1 The Human Timescale 2 Geological Timescale Millions of Years Periods Permian Triassic Jurassic Cretaceous Tertiary Quat. Major events Start of Age of Reptiles Dinosaurs Evolve and Expand
The Human Animal 1 The Human Timescale 2 Geological Timescale Millions of Years Periods Permian Triassic Jurassic Cretaceous Tertiary Quat. Major events Start of Age of Reptiles Dinosaurs Evolve and Expand
The Human Animal. Species. The Human Timescale. Geological Timescale. Primate Evolution Primate Ancestor
 The Human Animal The Human Timescale 1 2 Geological Timescale Species Millions of Years Periods Permian Triassic Jurassic Cretaceous Tertiary Quat. Major events Dinosaurs Evolve and Expand Start of Age
The Human Animal The Human Timescale 1 2 Geological Timescale Species Millions of Years Periods Permian Triassic Jurassic Cretaceous Tertiary Quat. Major events Dinosaurs Evolve and Expand Start of Age
8 Studying Hominids In ac t i v i t y 5, Using Fossil Evidence to Investigate Whale Evolution, you
 8 Studying Hominids In ac t i v i t y 5, Using Fossil Evidence to Investigate Whale Evolution, you were working with evidence for the evolution of the whale lineage. A lineage is a series of populations
8 Studying Hominids In ac t i v i t y 5, Using Fossil Evidence to Investigate Whale Evolution, you were working with evidence for the evolution of the whale lineage. A lineage is a series of populations
A new hominin from the Basal Member of the Hadar Formation, Dikika, Ethiopia, and its geological context
 Journal of Human Evolution 49 (2005) 499e514 A new hominin from the Basal Member of the Hadar Formation, Dikika, Ethiopia, and its geological context Zeresenay Alemseged a, *, Jonathan G. Wynn b, William
Journal of Human Evolution 49 (2005) 499e514 A new hominin from the Basal Member of the Hadar Formation, Dikika, Ethiopia, and its geological context Zeresenay Alemseged a, *, Jonathan G. Wynn b, William
Human Evolution Chris Stringer The Natural History Museum London. Are we nearly there yet?
 Human Evolution Chris Stringer The Natural History Museum London Are we nearly there yet? Phases of human evolution Human phase 2 0 Ma: >>Global spread Human anatomy >>Encephalised >>Dietary range >>Behavioural
Human Evolution Chris Stringer The Natural History Museum London Are we nearly there yet? Phases of human evolution Human phase 2 0 Ma: >>Global spread Human anatomy >>Encephalised >>Dietary range >>Behavioural
Overview of Hominin Evolution
 Overview of Hominin Evolution Lead Editor: Jessica Rothman, Katy Gonder, Holly Dunsworth, Kieran McNulty BIOLOGICAL ANTHROPOLOGY By: Herman Pontzer (Dept. of Anthropology, Hunter College; New York Consortium
Overview of Hominin Evolution Lead Editor: Jessica Rothman, Katy Gonder, Holly Dunsworth, Kieran McNulty BIOLOGICAL ANTHROPOLOGY By: Herman Pontzer (Dept. of Anthropology, Hunter College; New York Consortium
CHAPTER 9: HOMININ ORIGINS (PGS.
 Learning Objectives Explain the general time depth for the earliest primates and explain how they may (or not) be related to living primates Define what a hominin is and explain what sort of evidence is
Learning Objectives Explain the general time depth for the earliest primates and explain how they may (or not) be related to living primates Define what a hominin is and explain what sort of evidence is
Primate Evolution. Section 1. Primates
 Section 1 Primates Characteristics of Primates! Manual dexterity! Five digits on each hand and foot! Flat nails and sensitive areas on the ends of their digits! The first digits are opposable. Section
Section 1 Primates Characteristics of Primates! Manual dexterity! Five digits on each hand and foot! Flat nails and sensitive areas on the ends of their digits! The first digits are opposable. Section
Homework. Guided Reading Hominids Through Time (#12-21)
") Homework Guided Reading Hominids Through Time (#12-21) Learning Target I can explain how hominids evolved and what caused them to evolve. What characteristics do they have in common? What characteristics
Homework Guided Reading Hominids Through Time (#12-21) Learning Target I can explain how hominids evolved and what caused them to evolve. What characteristics do they have in common? What characteristics
Introduction to Biological Anthropology: Notes 17 The first hominins Copyright Bruce Owen 2008 Last time we saw how apes radiated (diversified) in
 in") Introduction to Biological Anthropology: Notes 17 The first hominins Copyright Bruce Owen 2008 Last time we saw how apes radiated (diversified) in the middle Miocene some shifted from quadrupedal to more
Introduction to Biological Anthropology: Notes 17 The first hominins Copyright Bruce Owen 2008 Last time we saw how apes radiated (diversified) in the middle Miocene some shifted from quadrupedal to more
Looking a fossil horse in the mouth! Using teeth to examine fossil horses!
 Looking a fossil horse in the mouth Using teeth to examine fossil horses Virginia Museum of Natural History Paleontology Department Fossil Teaching Kit 1 Teacher s Guide In this activity students will
Looking a fossil horse in the mouth Using teeth to examine fossil horses Virginia Museum of Natural History Paleontology Department Fossil Teaching Kit 1 Teacher s Guide In this activity students will
12/1/14. Speciation and Human Evolution. The Time Course of Speciation. Speciation Rates
 Speciation and Human Evolution References: chapters 24 (first few slides) 34 (last few pages of chapter) Speciation can occur rapidly or slowly, and can result from changes in few or many genes Many questions
Speciation and Human Evolution References: chapters 24 (first few slides) 34 (last few pages of chapter) Speciation can occur rapidly or slowly, and can result from changes in few or many genes Many questions
Chapter 17: Human Evolution
 Chapter 17: Human Evolution Classification Hierarchy Kingdom Phylum Class Order Family Genus Species Animal Chordate Mammal Primates Hominids Homo Sapiens Important Vocabulary Scientist who studies fossil
Chapter 17: Human Evolution Classification Hierarchy Kingdom Phylum Class Order Family Genus Species Animal Chordate Mammal Primates Hominids Homo Sapiens Important Vocabulary Scientist who studies fossil
Human Ancestry (Learning Objectives)
") Human Ancestry (Learning Objectives) 1. Identify the characters shared by all primates and relate them to the function they served in their common ancestor. 2. Learn the fields study of Human evolution
Human Ancestry (Learning Objectives) 1. Identify the characters shared by all primates and relate them to the function they served in their common ancestor. 2. Learn the fields study of Human evolution
Evolution-Human Evolution. Biology: Fezza Miami Arts Charter
 EvolutionHuman Evolution Biology: Fezza Miami Arts Charter Biogeography the study of the distribution of species and ecosystems in geographic space and through (geological) time Evolution is modification
EvolutionHuman Evolution Biology: Fezza Miami Arts Charter Biogeography the study of the distribution of species and ecosystems in geographic space and through (geological) time Evolution is modification
This is a repository copy of Middle Pliocene hominin diversity : Australopithecus deyiremeda and Kenyanthropus platyops.
 This is a repository copy of Middle Pliocene hominin diversity : Australopithecus deyiremeda and Kenyanthropus platyops. White Rose Research Online URL for this paper: http://eprints.whiterose.ac.uk/104209/
This is a repository copy of Middle Pliocene hominin diversity : Australopithecus deyiremeda and Kenyanthropus platyops. White Rose Research Online URL for this paper: http://eprints.whiterose.ac.uk/104209/
Anthro 101: Human Biological Evolution. Lecture 13: Early Hominins. Prof. Kenneth Feldmeier
 Anthro 101: Human Biological Evolution Lecture 13: Early Hominins Prof. Kenneth Feldmeier Biological Anthropology Hominoid = Apes Humans, Gorillas, Chimpanzees, Orangutans, Gibbons and Siamangs Hominin
Anthro 101: Human Biological Evolution Lecture 13: Early Hominins Prof. Kenneth Feldmeier Biological Anthropology Hominoid = Apes Humans, Gorillas, Chimpanzees, Orangutans, Gibbons and Siamangs Hominin
Lucy Redux: A Review of Research on Australopithecus afarensis
 YEARBOOK OF PHYSICAL ANTHROPOLOGY 52:2 48 (2009) Lucy Redux: A Review of Research on Australopithecus afarensis William H. Kimbel* and Lucas K. Delezene Institute of Human Origins, School of Human Evolution
YEARBOOK OF PHYSICAL ANTHROPOLOGY 52:2 48 (2009) Lucy Redux: A Review of Research on Australopithecus afarensis William H. Kimbel* and Lucas K. Delezene Institute of Human Origins, School of Human Evolution
Chapter 14: PRIMATE EVOLUTION
 Chapter 14: PRIMATE EVOLUTION PRIMATES What is a primate? Features that are unique to primates: -Present in primates -Absent in closely related groups Outgroup Ingroup Character A present Character A absent
Chapter 14: PRIMATE EVOLUTION PRIMATES What is a primate? Features that are unique to primates: -Present in primates -Absent in closely related groups Outgroup Ingroup Character A present Character A absent
Anthro 101: Human Biological Evolution. Lecture 13: Early Hominins. Prof. Kenneth Feldmeier
 Anthro 101: Human Biological Evolution Lecture 13: Early Hominins Prof. Kenneth Feldmeier Biological Anthropology Hominoid = Apes Humans, Gorillas, Chimpanzees, Orangutans, Gibbons Orangutan and Siamangs
Anthro 101: Human Biological Evolution Lecture 13: Early Hominins Prof. Kenneth Feldmeier Biological Anthropology Hominoid = Apes Humans, Gorillas, Chimpanzees, Orangutans, Gibbons Orangutan and Siamangs
Chapter 2 Human Origins: 7 Million to 1.9 Million Years Ago
 Chapter Overview Chapter 2 Human Origins: 7 Million to 1.9 Million Years Ago The chapter begins with a description of the Pleistocene epoch, which is also known as the Great Ice Age or the Ice Age. The
Chapter Overview Chapter 2 Human Origins: 7 Million to 1.9 Million Years Ago The chapter begins with a description of the Pleistocene epoch, which is also known as the Great Ice Age or the Ice Age. The
Anthro 101: Human Biological Evolution. Lecture 13: Early Hominins. Prof. Kenneth Feldmeier
 Anthro 101: Human Biological Evolution Lecture 13: Early Hominins Prof. Kenneth Feldmeier Biological Anthropology Hominoid = Apes Orangutan Humans, Gorillas, Chimpanzees, Orangutans, Gibbons and Siamangs
Anthro 101: Human Biological Evolution Lecture 13: Early Hominins Prof. Kenneth Feldmeier Biological Anthropology Hominoid = Apes Orangutan Humans, Gorillas, Chimpanzees, Orangutans, Gibbons and Siamangs
What do the Bones tell us?
 What do the Bones tell us? The scientific study of bones. Comes from the Greek word Osteon, meaning bone Sub-discipline of archaeology and physical anthropology, anatomy, forensics etc. Age at death Height/stature
What do the Bones tell us? The scientific study of bones. Comes from the Greek word Osteon, meaning bone Sub-discipline of archaeology and physical anthropology, anatomy, forensics etc. Age at death Height/stature
A New Kind of Ancestor: Ardipithecus Unveiled
 A New Kind of Ancestor: Ardipithecus Unveiled The oldest known hominin skeleton reveals the body plan of our very early ancestors and the upright origins of humankind Every day, scientists add new pages
A New Kind of Ancestor: Ardipithecus Unveiled The oldest known hominin skeleton reveals the body plan of our very early ancestors and the upright origins of humankind Every day, scientists add new pages
Build Vocabulary Students will have a more successful lab experience if they understand these terms.
 Guided Inquiry Forensics Lab hapter 26 Lab Investigating Hominoid Fossils Problem What can a comparison of skulls and hands reveal about the evolution of humans? Introduction paleontologist takes photographs
Guided Inquiry Forensics Lab hapter 26 Lab Investigating Hominoid Fossils Problem What can a comparison of skulls and hands reveal about the evolution of humans? Introduction paleontologist takes photographs
History matters: - personal basis - group basis
 Human Evolution History matters: - personal basis - group basis HISTORY GEOGRAPHY/CONTEXT humanity The recognition of the power of context and history motivates creationists Their concern: If we accept
Human Evolution History matters: - personal basis - group basis HISTORY GEOGRAPHY/CONTEXT humanity The recognition of the power of context and history motivates creationists Their concern: If we accept
The search for Adam's ancestors
 341 by Elaine Kennedy : 12 E volutionary biologists are convinced that humans are descendants of ape-like creatures. n spite of a number of disputes over theories of apehuman lineages, paleoanthropologists
341 by Elaine Kennedy : 12 E volutionary biologists are convinced that humans are descendants of ape-like creatures. n spite of a number of disputes over theories of apehuman lineages, paleoanthropologists
As we review the fossil evidence for early hominins, keep in mind the importance of identifying derived traits Ancestral traits are traits that have
 As we review the fossil evidence for early hominins, keep in mind the importance of identifying derived traits Ancestral traits are traits that have not changed from the earlier ancestral form Derived
As we review the fossil evidence for early hominins, keep in mind the importance of identifying derived traits Ancestral traits are traits that have not changed from the earlier ancestral form Derived
Hominins ultimately distinguished by brain size, bipedal locomotion and toolmaking behavior
 Early Hominins Hominins ultimately distinguished by brain size, bipedal locomotion and toolmaking behavior But these did not develop simultaneously: mosaic evolution The only reliable indicator of earliest
Early Hominins Hominins ultimately distinguished by brain size, bipedal locomotion and toolmaking behavior But these did not develop simultaneously: mosaic evolution The only reliable indicator of earliest
Introduction to Biological Anthropology: Notes 20 Apes and early hominins Copyright Bruce Owen 2010 the first known hominoids (apes) appeared in the
 appeared in the") Introduction to Biological Anthropology: Notes 20 Apes and early hominins Copyright Bruce Owen 2010 the first known hominoids (apes) appeared in the late Oligocene, 27 mya example Oligocene ape: genus
Introduction to Biological Anthropology: Notes 20 Apes and early hominins Copyright Bruce Owen 2010 the first known hominoids (apes) appeared in the late Oligocene, 27 mya example Oligocene ape: genus
BIO 182 LAB SIGN OFF PAGE LESSON 10
 BIO 182 LAB SIGN OFF PAGE LESSON 10 Name Please staple all of your lab pages for this Lesson together with this page as the top. You will use this page to get your Labs for Lesson 10 signed off by the
BIO 182 LAB SIGN OFF PAGE LESSON 10 Name Please staple all of your lab pages for this Lesson together with this page as the top. You will use this page to get your Labs for Lesson 10 signed off by the
(01) Ardipithecus kadabba The Smithsonian Institution's Human Origins Program
 Ardipithecus kadabba The Smithsonian Institution's Human Origins Program") (01) Ardipithecus kadabba The Smithsonian Institution's Human Origins Program Ardipithecus kadabba Where Lived: Eastern Africa (Middle Awash Valley, Ethiopia) When Lived: Between about 5.8 and 5.2 million
(01) Ardipithecus kadabba The Smithsonian Institution's Human Origins Program Ardipithecus kadabba Where Lived: Eastern Africa (Middle Awash Valley, Ethiopia) When Lived: Between about 5.8 and 5.2 million
Uncovering Ardipithecus Ramidus
 Uncovering Ardipithecus Ramidus Kristopher Jordan Krohn Mesa Community College/ Arizona State University 8 million years ago a tremendous even occurred; a new branch of primates split off from the chimpanzee
Uncovering Ardipithecus Ramidus Kristopher Jordan Krohn Mesa Community College/ Arizona State University 8 million years ago a tremendous even occurred; a new branch of primates split off from the chimpanzee
BIOL 1010 Introduction to Biology: The Evolution and Diversity of Life. Spring 2011 Sections A & B
 BIOL 1010 Introduction to Biology: The Evolution and Diversity of Life. Spring 2011 Sections A & B Steve Thompson: stthompson@valdosta.edu http://www.bioinfo4u.net 1 Human evolution where we came from
BIOL 1010 Introduction to Biology: The Evolution and Diversity of Life. Spring 2011 Sections A & B Steve Thompson: stthompson@valdosta.edu http://www.bioinfo4u.net 1 Human evolution where we came from
PATTERNS OF DENTAL SEXUAL DIMORPHISM IN EARLY HOMININS: A COMPARATIVE ACCOUNT
 Muséum national d Histoire naturelle Master Erasmus Mundus en Quaternaire et Préhistoire PATTERNS OF DENTAL SEXUAL DIMORPHISM IN EARLY HOMININS: A COMPARATIVE ACCOUNT ALIK HUSEYNOV Tuteur : PROF. DR. ROBERTO
Muséum national d Histoire naturelle Master Erasmus Mundus en Quaternaire et Préhistoire PATTERNS OF DENTAL SEXUAL DIMORPHISM IN EARLY HOMININS: A COMPARATIVE ACCOUNT ALIK HUSEYNOV Tuteur : PROF. DR. ROBERTO
The Origin and Evolution of Human Communication: If We Were Walking the Walk, Were We Walking the Talk?
 La Salle University La Salle University Digital Commons Explorer Café Explorer Connection 9-26-2018 The Origin and Evolution of Human Communication: If We Were Walking the Walk, Were We Walking the Talk?
La Salle University La Salle University Digital Commons Explorer Café Explorer Connection 9-26-2018 The Origin and Evolution of Human Communication: If We Were Walking the Walk, Were We Walking the Talk?
Knuckle-walking hominid ancestor: a reply to Corruccini & McHenry
 News and Views Brian G. Richmond David S. Strait Knuckle-walking hominid ancestor: a reply to Corruccini & McHenry Department of Anthropology, University of Illinois at Urbana-Champaign, Urbana, Illinois
News and Views Brian G. Richmond David S. Strait Knuckle-walking hominid ancestor: a reply to Corruccini & McHenry Department of Anthropology, University of Illinois at Urbana-Champaign, Urbana, Illinois
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature17654 Contents Supplementary Text S1. Calculating Locomotor Costs Supplementary Table 1. Estimated ranging costs for humans and other hominoids Supplementary Text S2. Estimating the Costs
doi:10.1038/nature17654 Contents Supplementary Text S1. Calculating Locomotor Costs Supplementary Table 1. Estimated ranging costs for humans and other hominoids Supplementary Text S2. Estimating the Costs
Hominid-Pongid Distinctiveness in the Miocene-Pliocene Fossil Record: The Lothagam Mandible
 AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 70:457-473 (1986) Hominid-Pongid Distinctiveness in the Miocene-Pliocene Fossil Record: The Lothagam Mandible ANDREW KRAMER Department of Anthropology, University
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 70:457-473 (1986) Hominid-Pongid Distinctiveness in the Miocene-Pliocene Fossil Record: The Lothagam Mandible ANDREW KRAMER Department of Anthropology, University
 2010-2014 www.d.umn.edu/cla/faculty/troufs/anthfood/aftexts.html#title 2010-2014 www.d.umn.edu/cla/faculty/troufs/anthfood/aftexts.html#title 2010-2014 www.d.umn.edu/cla/faculty/troufs/anthfood/aftexts.html#title
2010-2014 www.d.umn.edu/cla/faculty/troufs/anthfood/aftexts.html#title 2010-2014 www.d.umn.edu/cla/faculty/troufs/anthfood/aftexts.html#title 2010-2014 www.d.umn.edu/cla/faculty/troufs/anthfood/aftexts.html#title
Cotlow Award Application 2011
 Cotlow Award Application 2011 Department of Anthropology The George Washington University Washington, DC 20052 1. Personal Information Applicant s name: Heather Dingwall Degree sought: B.S., B.A. Field
Cotlow Award Application 2011 Department of Anthropology The George Washington University Washington, DC 20052 1. Personal Information Applicant s name: Heather Dingwall Degree sought: B.S., B.A. Field
Lecture Human Evolution
 Lecture Human Evolution I. Although modern human behavior is almost totally learned and cultural, it rests on a biological basis A. The processes of human evolution shaped humans brain and body 1. Accurate
Lecture Human Evolution I. Although modern human behavior is almost totally learned and cultural, it rests on a biological basis A. The processes of human evolution shaped humans brain and body 1. Accurate
AUSTRALOPITHECUS TO HOMO: Transformations in Body and Mind
 Annu. Rev. Anthropol. 2000. 29:125 46 Copyright c 2000 by Annual Reviews. All rights reserved AUSTRALOPITHECUS TO HOMO: Transformations in Body and Mind Henry M. McHenry and Katherine Coffing Department
Annu. Rev. Anthropol. 2000. 29:125 46 Copyright c 2000 by Annual Reviews. All rights reserved AUSTRALOPITHECUS TO HOMO: Transformations in Body and Mind Henry M. McHenry and Katherine Coffing Department
 2010-2014 www.d.umn.edu/cla/faculty/troufs/anthfood/aftexts.html#title 2010-2014 www.d.umn.edu/cla/faculty/troufs/anthfood/aftexts.html#title 2010-2014 www.d.umn.edu/cla/faculty/troufs/anthfood/aftexts.html#title
2010-2014 www.d.umn.edu/cla/faculty/troufs/anthfood/aftexts.html#title 2010-2014 www.d.umn.edu/cla/faculty/troufs/anthfood/aftexts.html#title 2010-2014 www.d.umn.edu/cla/faculty/troufs/anthfood/aftexts.html#title
Morphology of the Distal Radius in Extant Hominoids and Fossil Hominins: Implications for the Evolution of Bipedalism
 THE ANATOMICAL RECORD 295:454 464 (2012) Morphology of the Distal Radius in Extant Hominoids and Fossil Hominins: Implications for the Evolution of Bipedalism MELISSA TALLMAN* Department of Anthropology,
THE ANATOMICAL RECORD 295:454 464 (2012) Morphology of the Distal Radius in Extant Hominoids and Fossil Hominins: Implications for the Evolution of Bipedalism MELISSA TALLMAN* Department of Anthropology,
ROBUST AUSTRALOPITHECINES, OUR FAMILY TREE,
 Research TOC ROBUST AUSTRALOPITHECINES, OUR FAMILY TREE, AND HOMOPLASY Henry M. McHenry Darwin never predicted the existence of the robust australopithecines. 1 In his 1872 book The Descent of Man he foretold
Research TOC ROBUST AUSTRALOPITHECINES, OUR FAMILY TREE, AND HOMOPLASY Henry M. McHenry Darwin never predicted the existence of the robust australopithecines. 1 In his 1872 book The Descent of Man he foretold
THE EARLIEST HUMANS. Student Handouts, Inc.
 THE EARLIEST HUMANS Student Handouts, Inc. HOMINID EVOLUTION Hominids = great apes Chimpanzees, gorillas, humans, and orangutans Numerous intermediary fossils have been found But scientists disagree on
THE EARLIEST HUMANS Student Handouts, Inc. HOMINID EVOLUTION Hominids = great apes Chimpanzees, gorillas, humans, and orangutans Numerous intermediary fossils have been found But scientists disagree on
Project Description Form
 COTLOW FIELD RESEARCH FUND Department of Anthropology The George Washington University Washington, DC 20052 Project Description Form Applicant: Nicole L. Griffin Title of Project: Hominid Forefoot Kinematics,
COTLOW FIELD RESEARCH FUND Department of Anthropology The George Washington University Washington, DC 20052 Project Description Form Applicant: Nicole L. Griffin Title of Project: Hominid Forefoot Kinematics,
Short Film Great Transitions: The Origin of Humans IN-DEPTH FILM GUIDE
 DESCRIPTION IN-DEPTH FILM GUIDE Paleontologists have studied the fossil record of human evolution just as they have done for that of other major transitions including the transition from fish to tetrapods
DESCRIPTION IN-DEPTH FILM GUIDE Paleontologists have studied the fossil record of human evolution just as they have done for that of other major transitions including the transition from fish to tetrapods
Science Tear Sheet #4. Human Evolution: The Story, the Legend, and the Myth
 Science Tear Sheet #4. Human Evolution: The Story, the Legend, and the Myth The man who pleads his case first seems to be in the right; then his opponent comes and puts him to the test. Proverbs 18:17
Science Tear Sheet #4. Human Evolution: The Story, the Legend, and the Myth The man who pleads his case first seems to be in the right; then his opponent comes and puts him to the test. Proverbs 18:17
EARLY HUMANS COMPARE AND CONTRAST CHART
 Name: KEY Period: Date: World History Mrs. Schenck Early Human/ Nickname Ardipithecus ramidus Ardi Where they lived/ When Where: Eastern Africa (Ethiopia) When: 4.4 million years ago Very apelike, hairy
Name: KEY Period: Date: World History Mrs. Schenck Early Human/ Nickname Ardipithecus ramidus Ardi Where they lived/ When Where: Eastern Africa (Ethiopia) When: 4.4 million years ago Very apelike, hairy
Introduction to Biological Anthropology: Notes 21 Apes and early hominins Copyright Bruce Owen 2011 the first known hominoids (apes) appeared in the
 appeared in the") Introduction to Biological Anthropology: Notes 21 Apes and early hominins Copyright Bruce Owen 2011 the first known hominoids (apes) appeared in the late Oligocene, 27 mya example Oligocene ape: genus
Introduction to Biological Anthropology: Notes 21 Apes and early hominins Copyright Bruce Owen 2011 the first known hominoids (apes) appeared in the late Oligocene, 27 mya example Oligocene ape: genus
Journal of Human Evolution (1996) 30, Introduction
 30, Introduction") Lynne A. Isbell* Department of Anthropology, Rutgers University, Douglass Campus, Box 270, New Brunswick, New Jersey 08903, U.S.A. Truman P. Young Louis Calder Center, Fordham University, Box K, Armonk,
Lynne A. Isbell* Department of Anthropology, Rutgers University, Douglass Campus, Box 270, New Brunswick, New Jersey 08903, U.S.A. Truman P. Young Louis Calder Center, Fordham University, Box K, Armonk,
NEIL THOMAS ROACH. Division of Human Evolutionary Biology Harvard University
 NEIL THOMAS ROACH Division of Human Evolutionary Biology Harvard University 11 Divinity Avenue Email: ntroach@fas.harvard.edu Cambridge, MA 02138 Phone: 617-495-1870 EDUCATION 2012 PhD Harvard University,
NEIL THOMAS ROACH Division of Human Evolutionary Biology Harvard University 11 Divinity Avenue Email: ntroach@fas.harvard.edu Cambridge, MA 02138 Phone: 617-495-1870 EDUCATION 2012 PhD Harvard University,
NEIL THOMAS ROACH. Department of Human Evolutionary Biology Harvard University
 NEIL THOMAS ROACH Department of Human Evolutionary Biology Harvard University 11 Divinity Avenue Email: ntroach@fas.harvard.edu Cambridge, MA 02138 Phone: 617-495-1870 EDUCATION 2012 PhD Harvard University,
NEIL THOMAS ROACH Department of Human Evolutionary Biology Harvard University 11 Divinity Avenue Email: ntroach@fas.harvard.edu Cambridge, MA 02138 Phone: 617-495-1870 EDUCATION 2012 PhD Harvard University,
Adaptive Topographies, Sewall Wright s Shifting- Balance Theory and the Evolution of Horses.
 Adaptive Topographies, Sewall Wright s Shifting- Balance Theory and the Evolution of Horses. I. For a two alleles at a single locus, the change in gene frequency, p, from one generation to the next can
Adaptive Topographies, Sewall Wright s Shifting- Balance Theory and the Evolution of Horses. I. For a two alleles at a single locus, the change in gene frequency, p, from one generation to the next can
New fossil discoveries complicate the already devilish task of identifying HUMAN EVOLUTION Scientific American
 HUMAN EVOLUTION SHATTERED New fossil discoveries complicate the already devilish task of identifying 42 Scientific American, February 2013 ANCESTRY our most ancient progenitors By Katherine Harmon liest
HUMAN EVOLUTION SHATTERED New fossil discoveries complicate the already devilish task of identifying 42 Scientific American, February 2013 ANCESTRY our most ancient progenitors By Katherine Harmon liest
Journal of Human Evolution
 Journal of Human Evolution 64 (2013) 556e568 Contents lists available at SciVerse ScienceDirect Journal of Human Evolution journal homepage: www.elsevier.com/locate/jhevol Hominin stature, body mass, and
Journal of Human Evolution 64 (2013) 556e568 Contents lists available at SciVerse ScienceDirect Journal of Human Evolution journal homepage: www.elsevier.com/locate/jhevol Hominin stature, body mass, and
Comparing Indexes Among Primates
 CHAPTER 12 ADDITIONAL INVESTIGATION Comparing Indexes Among Primates Background Humans have the largest brains of all primates. In order to accommodate this large brain, the skull of a human has a vertical
CHAPTER 12 ADDITIONAL INVESTIGATION Comparing Indexes Among Primates Background Humans have the largest brains of all primates. In order to accommodate this large brain, the skull of a human has a vertical
b44 Australopithecenes < southern ape, root hominin, forest >
 149 b44 Australopithecenes < southern ape, root hominin, forest > We think back with repugnance to that ancient biological pre-human scene whence we came; there no life was a sacred thing. There, millions
149 b44 Australopithecenes < southern ape, root hominin, forest > We think back with repugnance to that ancient biological pre-human scene whence we came; there no life was a sacred thing. There, millions
ORIENTATION OF THE FORAMEN MAGNUM IN LATE MIOCENE TO EXTANT AFRICAN APES AND HOMINIDS
 Orientation of the Foramen Magnum in Late Miocene to Extant African Apes and Hominids XLIII/2 3 pp. 103 110 2005 MARTIN PICKFORD ORIENTATION OF THE FORAMEN MAGNUM IN LATE MIOCENE TO EXTANT AFRICAN APES
Orientation of the Foramen Magnum in Late Miocene to Extant African Apes and Hominids XLIII/2 3 pp. 103 110 2005 MARTIN PICKFORD ORIENTATION OF THE FORAMEN MAGNUM IN LATE MIOCENE TO EXTANT AFRICAN APES
Case Study: Climate, Biomes, and Equidae
 Case Study: Climate, Biomes, and Equidae In this activity, you will explore how environmental changes can shape life on Earth, using Equidae as a case study. By the end of the activity, you should be able
Case Study: Climate, Biomes, and Equidae In this activity, you will explore how environmental changes can shape life on Earth, using Equidae as a case study. By the end of the activity, you should be able
UDSM Researchers Discover 3.66 Million Years Old Hominid Footprints Frozen in the Ashes at Laetoli in Northern Tanzania
 UDSM Researchers Discover 3.66 Million Years Old Hominid Footprints Frozen in the Ashes at Laetoli in Northern Tanzania Summary Laetoli hominid footprints are the most persuasive evidence of habitual bipedal
UDSM Researchers Discover 3.66 Million Years Old Hominid Footprints Frozen in the Ashes at Laetoli in Northern Tanzania Summary Laetoli hominid footprints are the most persuasive evidence of habitual bipedal
Contribution to the knowledge of the fossil mammals from the ancient Tertiary terrains of Paraná. F. Ameghino * Tapiroidea
 Contribution to the knowledge of the fossil mammals from the ancient Tertiary terrains of Paraná F. Ameghino * Tapiroidea General characteristics: Upper molars with 2 transverse crests on the crown, each
Contribution to the knowledge of the fossil mammals from the ancient Tertiary terrains of Paraná F. Ameghino * Tapiroidea General characteristics: Upper molars with 2 transverse crests on the crown, each
REMEMBER YOU WILL NOT BE ABLE TO ANSWER THE QUESTIONS ABOUT ISLAND BIOGEOGRAPHY UNTIL AFTER THE 12/1 LECTURE
 REMEMBER YOU WILL NOT BE ABLE TO ANSWER THE QUESTIONS ABOUT ISLAND BIOGEOGRAPHY UNTIL AFTER THE 12/1 LECTURE Answers to Practice questions week 14 and 15 (Answers are in BOLD): 1) The above is the generally
REMEMBER YOU WILL NOT BE ABLE TO ANSWER THE QUESTIONS ABOUT ISLAND BIOGEOGRAPHY UNTIL AFTER THE 12/1 LECTURE Answers to Practice questions week 14 and 15 (Answers are in BOLD): 1) The above is the generally
Laetoli Footprints Preserve Earliest Direct Evidence of Human-Like Bipedal Biomechanics
 Laetoli Footprints Preserve Earliest Direct Evidence of Human-Like Bipedal Biomechanics David A. Raichlen 1 *, Adam D. Gordon 2, William E. H. Harcourt-Smith 3,4, Adam D. Foster 1, Wm. Randall Haas, Jr.
Laetoli Footprints Preserve Earliest Direct Evidence of Human-Like Bipedal Biomechanics David A. Raichlen 1 *, Adam D. Gordon 2, William E. H. Harcourt-Smith 3,4, Adam D. Foster 1, Wm. Randall Haas, Jr.
Analysis of performance at the 2007 Cricket World Cup
 Analysis of performance at the 2007 Cricket World Cup Petersen, C., Pyne, D.B., Portus, M.R., Cordy, J. and Dawson, B Cricket Australia, Department of Physiology, Australian Institute of Sport, Human Movement,
Analysis of performance at the 2007 Cricket World Cup Petersen, C., Pyne, D.B., Portus, M.R., Cordy, J. and Dawson, B Cricket Australia, Department of Physiology, Australian Institute of Sport, Human Movement,
The life history of Ardipithecus ramidus: a heterochronic model of sexual and social maturation
 Early origins of human sexual and social maturation Gary Clark, Maciej Henneberg Anthropological Review Vol. 78 (2), 109 132 (2015) The life history of Ardipithecus ramidus: a heterochronic model of sexual
Early origins of human sexual and social maturation Gary Clark, Maciej Henneberg Anthropological Review Vol. 78 (2), 109 132 (2015) The life history of Ardipithecus ramidus: a heterochronic model of sexual
Competitive Performance of Elite Olympic-Distance Triathletes: Reliability and Smallest Worthwhile Enhancement
 SPORTSCIENCE sportsci.org Original Research / Performance Competitive Performance of Elite Olympic-Distance Triathletes: Reliability and Smallest Worthwhile Enhancement Carl D Paton, Will G Hopkins Sportscience
SPORTSCIENCE sportsci.org Original Research / Performance Competitive Performance of Elite Olympic-Distance Triathletes: Reliability and Smallest Worthwhile Enhancement Carl D Paton, Will G Hopkins Sportscience
Session 16: Episode 5(1) Introducing Episode 5, our ancient ancestors and their relatives
 Introducing Episode 5, our ancient ancestors and their relatives") Session 16: Episode 5(1) Introducing Episode 5, our ancient ancestors and their relatives William P. Hall President Kororoit Institute Proponents and Supporters Assoc., Inc. - http://kororoit.org william-hall@bigpond.com
Session 16: Episode 5(1) Introducing Episode 5, our ancient ancestors and their relatives William P. Hall President Kororoit Institute Proponents and Supporters Assoc., Inc. - http://kororoit.org william-hall@bigpond.com
Sahelanthropus tchadensis: An Examination of its Hominin Affinities and Possible Phylogenetic Placement
 Totem: The University of Western Ontario Journal of Anthropology Volume 16 Issue 1 Article 5 6-21-2011 Sahelanthropus tchadensis: An Examination of its Hominin Affinities and Possible Phylogenetic Placement
Totem: The University of Western Ontario Journal of Anthropology Volume 16 Issue 1 Article 5 6-21-2011 Sahelanthropus tchadensis: An Examination of its Hominin Affinities and Possible Phylogenetic Placement
CHEIRIDIAL GENETICS AND TOOL USE AMONG LATER HOMINIDS
 XLVII/1 2 pp. 47 56 2009 DAVID WEBB, SARAH FABINY CHEIRIDIAL GENETICS AND TOOL USE AMONG LATER HOMINIDS ABSTRACT: There is still uncertainty about which species made the earliest known stone tools and
XLVII/1 2 pp. 47 56 2009 DAVID WEBB, SARAH FABINY CHEIRIDIAL GENETICS AND TOOL USE AMONG LATER HOMINIDS ABSTRACT: There is still uncertainty about which species made the earliest known stone tools and
Our own species, Homo sapiens, belongs to the order that also
 32 3 Primates and Human Origins Section 32 3 Our own species, Homo sapiens, belongs to the order that also includes lemurs, monkeys, and apes. Carolus Linnaeus named our order Primates, which means first
32 3 Primates and Human Origins Section 32 3 Our own species, Homo sapiens, belongs to the order that also includes lemurs, monkeys, and apes. Carolus Linnaeus named our order Primates, which means first
Cichlids of East Africa A Model of Vertebrate Radiation. ww.waveformenergetics.com
 Cichlids of East Africa A Model of Vertebrate Radiation ww.waveformenergetics.com www.wikipedia.com Lake Malawi 2-2020 million years old Fifth largest lake in the world by volume Bordered by Tanzania,
Cichlids of East Africa A Model of Vertebrate Radiation ww.waveformenergetics.com www.wikipedia.com Lake Malawi 2-2020 million years old Fifth largest lake in the world by volume Bordered by Tanzania,