The evolution of arthropodan locomotory mechanisms Part 10. Locomotory habits, morphology and evolution of the hexapod classes
|
|
|
- Samantha Owen
- 6 years ago
- Views:
Transcription
1 Zool. J. Linn. Soc., 51: With 6 plates and 40 figures AugustiNovember The evolution of arthropodan locomotory mechanisms Part 10. Locomotory habits, morphology and evolution of the hexapod classes S. M. MANTON (Mrs J. P. Harding), F.R.S., F.L.S. Zoology Department, British Museum (Natural History) and Queen Mary College, London Accepted for publication May 1971 An analysis has been made of the locarnotory movements and of the jumping escape reactions of certain hexapods, together with a description of their associated facilitating morphology. Common features shown by all hexapod gaits are due simply to the use of only three pairs of legs. Superimposed are features which are characteristic of each of the four apterygote classes and of the Pterygota. The associated morphology in each of these five classes is so distinct, and mutually exclusive, as to indicate a parallel evolution of the hexapodous state in the five classes, from multilegged animals with a common head type, differing from that of the Myriapoda. No one hexapod class could have been ancestral to any other. The morphological evolution of the Collembola has been associated with a hydrostatic jumping mechanism correlated with profound morphological peculirities. The jumping gaits and the jumping reactions implemented by the abdomen in Thysanura Machilidae are also correlated with unique morphology. A taxon, the Uniramia, is proposed to include the Onychcphora-M*apoda- Hexapoda. the Uniramia ranking as one of the several arthropodan phyla, and the Onychophora, Myriapoda and Hexapoda form threc subphyla of the phylum Uniramia. The conclusions are presented in but there is no separate summary. CONTENTS 1. Introduction. 2. Methods Terminology General characteristics of hexapod gaits A. Manner of stepping, fields of leg movement and tracks of hexapods B. The gaits of hexapods (i) The patterns of hexapod gaits (ii) The phase differences between successive legs.... (iii) The order of footfalls and the loading on the legs.. C. Conclusions concerning the general character of hexapod gaits 5. The running achievements of the Pterygota The locomotory achievements and habits of the apterygote hexapods A. Diplura (i) Habits and habitats of the Diplura (ii) Telescopic antenna1 movements of the Japygidae (iii) The runningachievementsof the Diplura..... (iv) Conclusions concerning dipluran locomotory achievements
2 204 S. M. MANTON B. Collembola (i) Habits and habitats of the Collembola (ii) The running abilities of Collembola (iii) The collembolan jump (iv) Conclusions concerning the collembolan running achievements... C.Thysanura (i) Habits and habitats of the Thysanura (ii) The walking gaits of Pepobius. (iii) The jumping gaits of Penobius (iv) The high jumpingescape reactionsofl'embius.... (v) The gaits of the Lepismatidae... (vi) Conclusions concerning the Thysanura D.Protura (i) Habits and habitats of the Protura (ii) The walking abilities of the Rotura (iii) Conclusions concerning the locomotory achievements of the Protura E. Conclusions concerning the locomotory achievements of the hexapod classes Thoracic pleuron. limb morphology and function in the apterygote classes... A. The pleuron. leg. leg-base and associated movements in the Diplura.... (i) The coxasternite articulation. its movements and the extrinsic leg muscles of the Diplura... Protractor and retractor muscles Levator and depressor muscles.. Depressor muscle from the trochanter (ii) Pleurites in the Diplura (iii) Dipluran legs. their intrinsic rocking mechanism and intrinsic muscles. The femoral rocking mechanism Intrinsic muscles B. The pleuron. leg. leg-base and associated movements in Collembola.... (i) The coxa-sternite junction. its movements and the extrinsic leg muscles in Collembola. Suspensory muscles.... Protractor and retractor muscles..... Abductor and adductor muscles Depressor muscle from the trochanter (ii) Pleurites and so-called subcoxae in Collembola (iii) Cod subdivisions and their functional and theoretical significance in Collembola (iv) Collembolan legs. their suspensory system and intrinsic muscles... Suspensorysystem Intrinsic muscles (for jumping mechanism and conclusions see $9 A. B) C. The pleuron. leg. leg-base and associated movements in the Thysanura... (i) Pleurites and their muscles in the Thysanura (ii) The coxa-pleurite junction and its movements in the Thysanura... (iii) Leg movements in the Thysanura (iv) Coxal musculature of the Thysanura Protractor and retractor muscles from the coxa Abductor and adductor muscles from the coxa (v) (vi) Other coxal muscles Leg structure and the special proficiencies of the Thysanura.... Morphology and movements of the coxa-trochanter joint... Muscles of the eoxa-trochanter joint Extrinsic levator muscles from the trochanter Muscles and joints of the telopod Summary of thysanuran peculiarities of thoracic and leg structure in association with habits (for abdomen and abdominal limbs see 0 9 C. D) D. The pleuron. leg. leg-base and associated movements in the Protura.... (i) Thoracic flexibility of the Protura (ii) The coxa-body articulation and its movements in the Protura.... (iii) The extrinsic rocking mechanism of proturan legs and leg structure.. E. Conclusions Concerning the pleuron and limbs of the apterygote classes... (i) The pleuron (ii) The leg and leg-base
3 EVOLUTION OF HEXAFOD LOCOMOTORY MECHANISMS 8. Thoracic endoskeleton and musculature A. Endoskeleton of the Diplura and Protura..... (i) Japygidae (ii) Campodeidae (iii) Conclusions concerning the endoskeleton of the Diplura and Protura. B. Dipluran thoracic musculature and its functional assets (i) Dorsal longitudinal muscles (ii) Ventral longitudinal muscles (iii) Deep dorso-ventral and deep oblique muscles (iv) Endoskeletal muscles (v) Superficial pleural muscles C. Conclusions concerning thoraclc InUSCUhture of the Diplura and Protura (for Collembola and Thysanura see 59 A(iv), (v). B. C) Trunk morphology and the jumping mechanisms of the apterygote classes... A. The collembolan jump (i) The cuticular connections and movements of the springing organ of Tomocerus (ii) The extrinsic muscles. and their functions. of the springing organ and hamula of Tomocerus (iii) The intrinsic muscles. their tendons and functions. of the springing organ of Tomocexus (iv) The trunk musculature of Tomocerus (v) The endoskeleton of Tomocerus Ribbon-like tendinous insertions The intersegmental tendons The paired endoskeletal plates (vi) Cuticular features of the collembolan trunk The cuticle The tergo-pleural arches B. Conclusions concerning collembolan structure (i) Cuticle and musculature (ii) Endoskeleton (iii) The structure of jumping and non-jumping Collembola..... C. The machilid jumping mechanism (i) Jumping (ii) Cuticular featuresof the abdomen ofperrobius (iii) The abdominal endoskeleton of Penobius (iv) The abdominal musculature of Perrobius D. Conclusions concerning the mode of action of the jumping mechanism of Petrobius General conclusions based on the analysis of hexapod movements. habits and associated morphology and the probable evolution of these classes. A. The evolution of the hexapod thorax B The evolution of hexapod gaits and speedy movement.... C. The evolution of external thoracic structure of the hexapod classes.... (i) Thoracic rigidity and flexibility (ii) The insertion of the legs..... (iii) Pleurites. subcoxal segments and flight D. The evolution of internal thoracic structure of the apterygote classes... (i) Endoskeleton of the thorax (ii) Extrinsic limb muscles of the thorax (iii) Thoracic trunk musculature E The evolution of thoracic leg StNCtuK in hexapods F The evolution of apterygote jumping G. The evolution of hexapod classes Acknowledgements References Appendix 1. The muscular system of the trunk ot the jumping collembolan Tomocerus longicornis (i) The dorsal longitudinal system (ii) The lateral longitudinal system (iii) The sternal longitudinal system (iv) Thedeep dorso-ventral anddeep obliquemuscles (v) The superficial pleural muscles (vi) The muscles of the hamula

4 206 S. M. MANTON (vii) The muscles of the ventral tube (viii) Suspensory muscles from the coxa to the endoskeleton 394 Appendix 11. The cuticular connections of the thoracic endoskeletal system of Tomocerus longicornis INTRODUCTION An investigation of the modes of action of the mandibles and mouth parts of Symphyla and other Myriapoda (Manton, 1964) and the study of the functional morphology associated with the locomotory habits of these animals (Part 9 and previous Parts of the present series ), has shown how very different are these systems from those of pterygote insects, differences so deep-seated as to preclude any close relationship between the Myriapoda and Pterygota. Among the apterygote classes the parallel evolution of entognathy has been demonstrated (Manton, 1964). It now remains to show the functional significance of the trunk morphology of the apterygotes and to determine, in respect of locomotory habits, what evidence concerning their evolution can be obtained from this source. Of particular importance is a study of the pleuron and leg-base, because the apterygote classes have been supposed to illustrate certain basic conditions leading towards those of the Pterygota. But even the morphology of the apterygotes has not been known with accuracy. And there has been no understanding concerning the functional significance of pleural structure, without which there can be no proper appreciation of the paths of evolution. The results of such a study, presented here, shows that the apterygote classes are fundamentally different from one another. Summaries are given in the concluding paragraphs of many sections, in addition to those marked Conclusions. 2. METHODS The methods employed are similar to those described in previous Parts but with certain additions described below. In the many-legged Myriapoda it is usually possible to determine the pattern? of a gait and the phase difference between successive legs from still photographs, combined with records of the footprints. Such determinations of the gait can be made much more speedily than by the exclusive analysis of cinematograph films. But still-photographs of hexapods are not readily interpreted in the same way. Many films of hexapods taken at normal frequency have been analysed, but gait determination by &is means is limited, because in the fast stepping species there may be only two or three frames per cycle of leg movements, and sometimes even less as in the jumping gaits of Petrobius, which show only one frame per cycle during fast progression. When problems have been great, still photography of various kinds has provided the answers. * References in the text to Parts 1-9 of this series will be given as Part 1, etc. without date (for details see list of references). t The pattern of a gait means the relative duration of forward to backward strokes, a term employed in previous Parts.
5 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 207 The labour of analysing great lengths of high speed or normal cinematograph films in order to ascertain the pattern of a gait can easily be avoided. Some 40 to 50 good quality still photographs of a running hexapod usually show the range in positions of the legs. The fields of movement of the limbs, such as shown in figures 51, 6m can thereby be ascertained. When the extreme forward and backward positions of the legs in contact with the ground are known, it is possible to judge how far through the backstroke any particular legs on one photograph may be. Sometimes it is not easy to be sure whether, in a photograph, a leg is on or off the ground. A shadow meeting, or not meeting, the limb tip may be decisive (pls. 1, figures a, h; 6, figures e, f). The forward swing of a hexapod leg is usually executed pore rapidly than the backstroke, and the tip of such a limb moves forwards over the ground more rapidly than the body. An exposure time which gives a sharp image of the body and of some limbs, while others appear blurred, differentiates the propulsive limbs, with tips on the ground, from the forwardly moving limbs, which appear blurred* (pls. 1, figures f, m; 5, figures a, h, g). And the taking of a still photograph of a hexapod which has just run off smoked and on to white paper, so that a good picture is obtained of the limbs as well as the track, as in pl. 1, figure a, provides the stride length and the immediate past positions of the legs on the ground. This simple practice is useful when the legs are dark in colour and do not show up well over the soot paper, pale coloured legs are quite distinct against a black background (pls. 1, figure c; 2, figure b; 3, figuree; 5, figures a, c, d). Lastly, limb tips usually do not slip on the ground, and consequently lie at the same distance from the middle of the track throughout the backstroke. But during the forward stroke the limb tip off the ground extends further out from the body. Measurement of the distance between the limb tip and middle line of the track is sometimes useful in determining whether a limb in a photo is on or off the ground. Two types of diagram have been used in determining the characteristics of a gait from photographs showing which legs are on and off the ground. Diagrams such as figures 4a-e, 5g-i, etc., which show the positions of the legs along the body at one moment in time, will be referred to as gait stills. Diagrams such as figures 2a-c, 3a-c, etc. are gait diagrams which present each leg in recovery and propulsion (the pattern), to scale and successive and paired legs are in their correct phase. A horizontal run is a time interval, and therefore a vertical line, such as indicated by short arrowed lines on figure 3a, b, represents an instant in time. The actual positions of the legs along the body at this instant can be extrapolated from the vertical line, so forming a gait still diagram. The next step is to select a photograph showing right leg 3 at the end of the backstroke and to match it with a fuller series of gait stills than those on figure 4a-e. This will give an approximation towards the gait. An exact determination of the gait can then be made by reference to a series of gait diagrams, such as those on figures 2, 3, 5-7, ascertaining first whether a vertical line through the position of right leg 3 at the end of the backstroke will lead to a predictable gait still which corresponds with the photograph. If the other five legs do not exactly fit, then the changes required in the pattern of the gait and * The depth of focus is of course adequate to give no blurred images of forwardly moving limbs at shorter exposure times.
6 208 S. M. MANTON phase difference between successive legs can be ascertained so that the gait diagram fits this and other photographs. There is some momentary digression from mathematical accuracy in stepping by the animals, but on the whole their stepping is remarkably regular, especially when running fast. The range of gaits, or at any rate the faster end of the series, can also be ascertained from a number of still photographs, particularly if they are taken under different conditions and at different times. A small number of photographs of the same hexapod taken in rapid succession usually show the same gait. Under bright photographic lights most hexapods use their faster patterns of gait. The recorded speeds refer to experimental runs at room temperature by freshly caught hexapods. The temperature has an effect on speeds, as in other cold-blooded animals. In obtaining evidence from photographs of the presence of a leg-rocking mechanism, which brings the dorsal surface forwards during the propulsive backstroke and backwards during the forward stroke, so assisting the propulsive extension of leg joints lacking extensor muscles (see Part 8, figures 2, 3 and p. 258), the position of the axis of swing of the coxa on the body must be known. When this axis is approximately vertical (Part 6, text-fig. 21a, d) the leg will show its dorsal surface from above or ventral surface from below at all moments during the propulsive backstroke (as in Part 4, pl. 54, figure 28; Part 6, pl. 9, figures 28, 29). When the axis of coxal swing on the body is approximately horizontal (Part 6, text-figure 4a, b) the anterior face of the leg will be visible in dorsal view at the start of the backstroke and the posterior face will be visible during the latter part of the backstroke, when the leg swings approximately equally fore and aft of its base. This movement is shown fairly clearly in Part4, pls. 53, figure 22; 55, figure 38 and less clearly in pl. 53, figure 25. The effect upon the leg of these two contrasting axes of swing of the coxa are most easily apprehended from working models (see Part 6, text-fig. 3e, f). When the coxa possesses a vertical axis of swing, and, unlike the example given above, the anterior, dorsal and posterior faces of the leg are visible in turn in dorsal view, a rocking mechanism must be present (Part 8, pl. 2, figures 107, 108, 110, described on p. 258). When the axis of swing is horizontal, no separate rocking mechanism is needed to assist leg extension because a rock occurs automatically. The importance of the comparative approach to functional morphology cannot be overestimated. An essential tool throughout the present work has been the knowledge already obtained concerning arthropod structure and how it functions. Without this information the significance of collembolan structure, for example, might not have been unravelled so readily. 3. TERMINOLOGY The term sclerite or scute designates a local sclerotization of the cuticle forming a stiffened zone. Throughout Parts 2-9 the word sclerite is used in this sense. A definite margin to a sclerite may be present, or the margin may merge structurally into the flexible surrounding cuticle (Part 8, figures 6-15). The total complement of sclerites may invest the body completely, as in most Diplopoda (Part 7, text-figs. 1, 4, 11) and Geophilomorpha (Part 8, figures 41, 45), or it may not. When the sclerites are separated by flexible
7 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 209 cuticle which is little sclerotized, as in Symphyla (Part 9, figures 5, 6), most Chilopoda (Part 8, figures 1,48, 58) and the apterygote classes considered below, the pleural names are limited here to the sclerites themselves. To state that a certain pleurite is entirely membranous, as in descriptive entomology, is considered here to be a contradiction in terms. Tergites and sternites are roughly comparable in all classes. It appears to be reasonable in some cases to call the lateral parts of a dorsal sclerite a paratergite, whether it is separated or not, but in others the lateral part of a large dorsal sclerite is clearly a pleurite undivided from the tergite. Snodgrass (1960) objects to the use of these long established terms in favour of tergum and sternum. But tergum, the back L., is not always armed with a tergite. There are no tergites on the terga of the thorax of a barnacle or on the abdomen of a hermit crab. Similarly there are no sternites on the thoracic sterna of Collembola and Machilidae, while sternites are well developed here in Diplura and Lepismatidae. Both terga and sterna sometimes carry more than one sclerite. These terms have been in use for a century by writers on all classes of arthropods, together with the customary adjectives, tergal and sternal, and should stand. The pleuron. It appears to be customary in some entomological literature to map the whole pleuron into areas bearing the names of sclerites, regardless of their degree of sclerotization. These sclerites are further equated with metamorphosed pieces of a theoretical subcoxal ring, a leg segment which is supposed to have been incorporated into the pleuron. Neither of these practices finds support here. The pleurites show much variation in number, in position, in detailed structure and in function. There is no reason why the pleural sclerites of all classes should be regarded as modifications of a common basic plan. In hexapods, as in myriapods, pleurites may differ on consecutive segments, as in most Scolopendromorpha and Craterostigmus (Part 8, figures 1, 74, 75) and see the pro- and mesothorax of Petrobius (figures 31b, 32 and A($, B, C, 10 C(iii)). The erection of hypothetical basic pleural morphology and the deriving of the morphology of extant classes from it, seems to be a zoological pastime which brings no fundamental understanding, because the hypothetical basic form of the pleurites has in no way been related to function. An arthropod, as well as any other animal, must have been a working concern at all stages in its evolution. Pleural morphology is flexible, and function determines the number and form of the pleurites on any one segment. The terms anapleure and katapleure as applied to hexapods, does not indicate a homology between the sclerites so named in the Myriapoda. But in order to avoid confusion with the entomological literature, the pleurites of the apterygote classes are refered to below either by numbers or.by their customary entomological names, but with no implication of homology with the similarly named sclerites in other classes. The terms promotor and remotor refer to the whole of the stepping movement of a leg; the former indicates the forward swing, off the ground, implemented by protractor, levator, sometimes flexor and other muscles, while the latter indicates the backward power stroke of the leg with its tip in contact with the ground, implemented by retractor, depressor, sometimes flexor and other muscles.

8 210 S. M. MANTON The terms walking and running roughly refer to speeds of progression in most animals, but when applied to the many-legged invertebrates do not have precisely the same meaning as for bipedal or quadrupedal vertebrates. In walking, one foot of a pair is always on the ground, while running gives swifter progression and shows both feet off the ground for a proportion of a pace. On these definitions most hexapods, however fast they travel, do not run but walk, while myriapods run or walk according to the gaits employed. Professor J. B. S. Haldane (1964, pers. comm.) defined walking and running in another manner: We say that a man walks if the period of his steps is about equal to that of his legs considered as pendulums, and runs if it is less. There is no word for slower speeds. I will employ the word slouching. I take it that walking involves less waste of energy than running or slouching. Clearly for any arthropod such a classification is a pure formality, because the resistance of the tissues, and of water or even air, would presumably damp pendulum movement pretty quickly. Formally however, I should guess that all arthropods slouch.... For a geometrically similar series of animals the walking step length must vary as the square root of the linear dimension. It is just about right for man.... Man is relatively efficient in a way not yet noticed, and his dimensions are nearly optimal (of course other reasons against being 12 or 3 feet high are more important). Since the distinction between walking, running or slouching in myriapods and hexapods is of little value in appreciating the differences between their several gaits, the words walking and running will be used below in their colloquial sense, implying slower and faster progression. The unique jumping or hopping gaits of the Thysanura are defined in their description (0 0 6 C(iii), 7 C(iii)). They share some features with quadrupedal galloping and bipedal jumping and hopping in vertebrates. 4. GENERAL CHARACTERISTICS OF HEXAPOD GAITS An analysis of the gaits of a number of myriapods and of hexapods was made after those of the Onychophora were recorded in Part 1, and some comparative features of these gaits were summarized in Part 2. It was clear from this work that a proper understanding of hexapodous running, of the associated functional significance of differing class morphologies and of the pre-requisites for the evolution of flight, would only be possible after the work recorded in Parts 3-9, and the functional morphology presented below, was completed. Both before and since the publication of Part 2 several descriptive accounts of insect gaits have been published (for references see Hughes, 1965). 4A. MANNER OF STEPPING, FIELDS OF LEG MOVEMENT AND TRACKS OF HEXAPODS There is no doubt that hexapodous states have been evolved from animals in which more than three pairs of legs were locomotory (Part 2). The advantages of using only three pairs of legs are several. Increase in leg length can give increase in speed of running because longer strides can be used, but long legs in a many-legged animal leads to difficulties because the gaits must be executed
9 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 21 1 very exactly if interference with one leg by another is to be avoided (Ligia, Scutigeru, Part 2, text-figs. 2, 3). A fanning-out of the anterior and posterior legs reduces the overlap of the fields of movement of successive legs, and when only three pairs are used, the fields scarcely overlap at all. The fields of movement of the legs of Gmpodea, Tomocerus and Petrobius are shown in figures 5j, 6m, 7a and of Forficulu in Part 2, text-fig. 3. A second advantage in the non-overlap of fields of movement is the ease with which the gaits can be changed. The overlapping fields of Scutigeru and Ligia result in little change in the practicable relative durations of the forward- and backstrokes (see Part 2, Table 2, column 1). Increase in speed of running in most arthropods is achieved, not only by the use of faster patterns of gait, but by decrease in the pace duration and increase in the angle of swing of the leg (see Part 2, Table 1). The stepping action of leg 1 of a hexapod may take place in front or largely in front of the origin of this leg Erom the body, and that of leg 3 usually takes place well behind the base of this leg. In the former the propulsive backstroke of the leg against the ground is largely one of progressive flexure while in the latter it is one of progressive extension. Leg 2, or any other leg in which the field of movement lies both in front and behind the base of the leg, performs a propulsive backstroke in which progressive flexure takes place first, followed by propulsive extension, as in most myriapods. Since all legs must take the same stride length, legs 1 in hexapods move through a larger angle of swing than the longer legs 3. It is necessary for the three pairs of legs of a hexapod to be closely based, it would not do to have one anterior pair, one in the middle, and one at the end of the body. Legs of each pair usually need to work in opposite phase, and if widely spaced the leverage they would exert would throw the body into undesirable undulations. The three pairs of legs need to be on successive segments, and they need to be close to the head to give support to feeding movements. Many hexapods living in soil, decaying logs, etc., have such short legs t5at most of the above-mentioned advantages of the hexapodous state do not obtain, e.g. some Collembola and japygids, pls. 2, figures d-i; 3 figures h, i. Many sources of evidence suggest that these short legs are secondary and confer on their owners the ability to penetrate easily into minute spaces, a habit of survival value in face of an ever growing efficiency in carnivorous surface running hexapods, many of which can also fly, and in other predators. Since hexapodous stepping has been evolved independently many times among widely different arthropodan classes (examples from Crustacea and Arachnida were given in Part 2, text-figs. 2, 3), it is possible that this stepping may have been acquired independently by the several hexapod classes. There is only a limited number of ways in which hexapodous stepping can take place, and therefore some similarities between the hexapods may be expected to be due to convergence (see further, 5 10 G). When only three pairs of legs are present it is usually necessary for three legs to be in contact with the ground at all moments throughout the pace in order that stability be maintained, rhe centre of gravity being situated within the triangle between these limb tip. It is desirable for leg n + I to be put on the ground just before leg n is raised, or simultaneously with this event, so that the
10 212 S. M. MANTON body is not left momentarily unsupported by two successive legs. In the Onychophora the leg is blunt and wide at its extremity, leg n + 1 is put on the ground after leg n is raised, support for the body being given by more remote limbs, and a suitable angle of swing of the leg is thereby accomplished. A narrow pointed limb tip enables leg n +I to be put on the ground simultaneously with, or in advance of, the raising of leg n without causing mechanical interference, contrast the stepping shown in Part 1, text-fig. 6a, b, with those given here in figures 2a-c, 3a-c, 5b, 7b, c, the time interval k indicating the period during which the two successive legs, occupying the same group of footprints, are both supporting the body (see also Part 2, p. 102). The possession of a narrow pointed limb tip promotes a long stride and faster walking than shown by Onychophora, and makes the hexapod condition mechanically possible. When a narrow-jointed tarsus is plantigrade, as in the Pterygota, leg 2 is placed upon the ground lateral to the tarsus of leg 1 before this leg is raised, and at almost the same transverse level as the tip of leg 1, or close behind it (see right legs on pl. 1, (figures a, c). Leg 3 is put on the ground close behind and median to leg tip 2 (see pl. 1, figure a where left leg 3 is about to be put on the ground, and pl. 1, figure e where the footfall of this leg has taken place). This arrangement of the footprints is seen on the tracks, pl. 1, figures a, b, c, although the stride length (distance apart of the groups of footprints of one side) differs in (a) and (b), (see also the diagrams on figure lf, g). Where the stance is unguligrade, as in the apterygote classes, then the narrow pointed tarsus of leg 2 can, if this leg is considerably longer than leg 1, be put on the ground outside and in front of the tip of leg 1 without causing mechanical interference, see Cumpodeu, pl. 2, figures a, b where right leg 2 is about to be put on the ground.* Leg 3 can also be put on the ground outside and in front of leg 2 before leg 2 is raised, see left leg 3 p1.2, figures a, c. In this manner the three footprints in each group on the track are forwardly staggered, that of leg 3 being the most anterior and lateral of the group, instead of being the most posterior, as in Pterygota (see figure lb). When pointed unguligrade legs are not so long and do not show such marked differential length along the body, as in Jupyx, leg 2 is placed on the ground behind leg 1 and leg 3 behind leg 2, so forming groups of footprints staggered in the opposite direction to those of Cizmpodeu (fpre lc). In the Collembola the limbs are unguligrade but the footfalls are placed somewhat as in the Pterygota, the differences in leg length on successive segments being much smaller than in Cumpodeu (see pl. 3, figure e where left leg 3 is at the end of the backstroke, its tip being in between the footprints of legs 1 and 2 while right leg 3 is about to be put on the ground in this position, just before leg 2 is raised). When the legs of a pair are used in opposite phase, as is usual in hexapods (Part 2, p. 111), the groups of footprints on the two sides of the track alternate, as is common in hexapods. Rut when the legs of each pair are moved in similar phase, the footprints on the two sides of the track are level with each other. A rough comparison is shown in figure 1 between representative tracks Whether a leg tip is on the ground or off it in the photographs has been carefully determined by analysis of the gait, stride lengths and tracks. Where a shadow of a leg meets the legtip, then the latter is on the ground, when a shadow does not meet the limb tip, see pi. 6, fwra e, f, the leg is off the ground but shadows are not always present.
11 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 21 3 CAMFQDEA TOMOCERUS JAPYX 23 I a 132 b THERMOBIA 3 2 PETROBIUS I d e l I f 21 3 BEETLE HELOPS I2 3 2l I l HEMIPTERAN 1 3 I Figure 1. Diagram showing footprint positions, from tracks made by hexapods running over smoked paper and performing their fastest gaits. The tracks are scaled to a roughly common thoracic segment volume in order to show comparisons between species. Actual tracks are seen on pls The median line between the right and left footprints measures two stride lengths and the numbers 1-3 indicate the positions of the footprints of the three thoracic legs. (a) Collembola, Tomocems Zongicomis Muller. (b-c) Diplura, (b) campodeid, (c) japygid. (d-e) Thysanura, (d) Thermobia sp., (e) Peeobius brevirvlis Carpenter. (f-g) Pterygota, (f) Helops luevioctostriatus Gaaa (Coleoptera), (g) DoZycoris baccarum (L.) (Hemiptera).

12 214 S. M. MANTON of the several groups of hexapods, based upon a calculated common segment volume, this giving a measure of comparison between the leg and the stride lengths of the animals shown. The vertical lines in the middle of each track represents the length of two successive strides. The tracks (figure 1) of two pterygotes, a beetle and a hemipteran, are wide because the legs are long, but the stride lengths are not large, indicating that either very small angles of swing of the leg, or slow patterns of gait, were used. Greater variety is shown by the tracks of the Collembola and Diplura on figure la, b, c. In Japvx the track is narrow because the thoracic segments are narrower and longer than those of most Pterygota. The footprints in each group are backwardly staggered because the legs are short, but the strides are longer than those of the two pterygotes shown. In the other dipluran, Campodea, the groups of footprints are forwardly staggered (see above), the stride is very long, as are the legs, which also show large differences in length along the thorax. In the collembolan Tomocerus the track is much as in the Pterygota, except that the unguligrade stance and narrow limb tips enable the footfalls in each group to lies at the same transverse levels. In the Thysanura (figure Id, e) Thermobia, with very much shorter legs than Petrobius, shows groups of forwardly staggered footprints remarkably close in to the middle line (cf. Pterygota), and a short stride. The jumping gait of Petrobius is given on the right in which, unlike all the other tracks shown, the groups of footprints from the two sides of the body are level with each other, each pair of legs being moved in the same phase. The stride is longer than in Thermobia. The walking gait of Petrobius, used on the under side of rocks, etc. resembles those of other hexapods in the paired legs being moved in opposite phase ( Q 6 C(ii)). Conclusions. The differences in stepping of the hexapod classes, indicated by the positions of their footprints on the tracks, are considerable. The various types of track are controlled by the length of leg, both in respect of body size and relative to the other legs along the thorax; by the pattern of the gait and phase difference between legs of a pair; and by the presence or absence of a plantigrade stance. Only the Pterygota possess the latter, although not always in larval stages (pl. 2, figures g, h), and their tracks contrast with those of the unguligrade, apterygote classes. But the tracks of the latter, although less distinct from one another, show their class characteristics. The distinctions are correlated with the different mechanisms and morphology of the leg base (8 7). These features are mutually exclusive from class to class (see below), thus indicating that the general similarities in fields of movement and of tracks e.g. figure lb, d, are convergent, there being only a limited number of ways in which three pairs of legs can be moved, and that the differences imply parallel evolution of hexapodous states (see further, 0 10 G). 4B. THE GAITS OF HEXAPODS As in previous Parts, the relative durations of the forward to the backward strokes of the legs are refered to as the pattern of the gait and are expressed by numbers within brackets, whose sum is 10. The phase difference between successive legs, p.d., is expressed as that proportion of a pace by which leg n + I is in advance of leg n. The term gait includes both the relative durations
13 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 215 of forward and backward strokes and the phase difference between successive legs and is illustrated by the gait diagrams on the figures. (i) The patterns of hexapod gaits It has been noted in Part 2 and by von Holst (1934), Hughes (1952, etc.) that for stability to be maintained at all moments during running, a hexapod must always have at least three legs in contact with the ground, disposed around the centre of gravity; the paired legs must move in opposite phase, except when swimming or jumping; and leg 1 must remain on the ground until the footfall of leg 2 on the same side of the body, and similarly for legs 2 and 3. It is shown below that these generalizations do not apply to all apterygote classes. The gait patterns which meet the above considerations are limited; the backstroke can never be relatively shorter than the forward stroke if stability is to be maintained at all times. The pattern (5.0 : 5.0) can change in the slower direction towards (1.5 : 8.5), but if the backstroke becomes the shorter in relative duration, as in gait (5.5 : 4.5) then there will be momentaryperiods of instability with only two legs in contact with the ground. The fast patterns of gait employed by the myriapod classes in achieving speed are impracticable to a hexapod; the fastest scolopendromorph gait is about (8.5 : 1.5), showing only three out of forty legs in full propulsive phase at any one moment (Part 8, figure 28d, Part 3, text-fig. 3), the fastest lithobiomorph and scutigeromorph gaits are about (6.5 : 3.5) Part 3, text-fs. 6, the fastest diplopod gait is about (6.4 : 3.6), Part 6, Table 1, p. 495, and the fastest pauropod gait is about (7.5 : 2.5), Part 3, text-fig. 12. A hexapod can show considerable range in the pattern of its gaits, but not the same in each species, and most patterns are medium to slow. Changes in the pattern of gait produce changes in the speeds of running, but the contributions by gait change are small compared with those caused by changes in the pace duration and in the angle of swing of the leg, but the contribution is a significant one, see entry for the earwig in Part 2, Table 1, p (ii) The phase difference between successive legs The phase difference between successive legs, p.d., is defined in the previous section. As in myriapods the p.d. of hexapods is dependent upon the pattern of the gait for mechanical reasons and changes with it. The hexapod p.d. is <0.5 in the usual stable gaits, but becomes >0.5 in the few hexapods which use unstable gaits (fast gaits of Campodea and Thysanura). The effects of altering a phase difference between successive legs from 0.33 to 0.25, while the pattern of the gait remains the same at (2.0 : 8.0), is shown in figure 2a-c. The time interval k, during which two successive legs are in contact with the ground and occupy the same group of footprints, is marked on (b). A reduction in phase difference between successive legs shortens this time interval, as in (c), and an increase lengthens it, as in (a), the pattern of gait remaining the same. For this pattern, the phase difference between successive legs cannot be less than 0.2, or the interval k becomes non-existent, and an unsupported moment ensues at the change of successively supporting legs, giving instability. When the fraction of
14 216 S. M. MANTON the pace spent by each leg off the ground is equal to the fraction it is out of phase with the next leg, shown by the number for the relative duration of the backstroke being equal to that for the phase difference between successive legs (omitting the difference of ten used here to differentiate the one from the other), then the time interval k, as on figure 3b, will cease to exist and Legs n and n + I will be picked up and put down on the ground simultaneously, and the stability conferred by the time interval k will be eliminated. Whether the period k is long, short, or absent depends upon the species and the speed of running. A long period k is recorded by Hughes (1952, text-fig. 5 B, C), a very short one on his text-fig. 7 for Bhtta, and irregular long and short periods k for Dytiscus on his text-fig. 5D. An almost simultaneous change over is shown by many hexapods, either at all patterns of gait, or only at the fastest speeds. The heavier pterygotes and the more slowly moving species show substantial time intervals k giving stability, while among the apterygotes the time interval k may be very short or non-existent, as on figures 5a and 6. (iii) The order of footfalls and the loading on the legs Roeder (1937) noted that the rhythm of leg movements, indicated by the order of the footfalls, could vary according to whether the insect was walking quietly, or was more excited, or climbing. Hughes (1952) records further variations in the order of footfalls during each pace. The way in which the various orders of footfalls, and the time intervals between them, are controlled by the pattern of the gait and the phase difference between successive legs, is readily seen in figures 2-4. On figure 2 the order of footfalls (d-f) is shown below the three gait diagrams (a-c) and the number of propulsive legs at moments throughout the pace in (g-i). In gait (a) the interval between the footfdls are equal (d) and there are always five legs in the propulsive phase (g). Even loading on the legs results. In (e) and (f) the footfalls of legs 3 and 2 are retarded relative to the footfalls of legs 1 on the same side (they shift to the right along the pace line), until footfalls of legs 1 and 3 on opposite sides are simultaneous (f). In gait (b) successive footfalls are unevenly distributed and the propulsive legs at any one moment range from four to five in number (h). In gait (c), where the footfalls of legs 1 and 3 on opposite sides of the body are simultaneous (f), the supporting legs range from four to six in number (i). The most uneven loading on the legs occurs in gait (c). Gait (a), with even loading on the legs, is frequently seen on photographic records of hexapods, and the moments in time shown on the plates are marked by arrows on the gait diagrams. Thus the hexapods often exhibit preference for even loading on the legs and even footfall intervals, as do the Chilopoda (Part 3, p. 143). The pattern of the gait with even loading (2.0 : 8.0) is a slow one, although possessing mechanical advantages. Increase in speed can result from a reduction in the relative duration of the backstroke, but the advantages of even loading on the legs, shown by figure 2a, g, cannot be maintained. Uneven loading on the legs becomes inevitable, but it can be minimized by selection of the least unfavourable gaits which give faster running. A relative duration of forward to backstroke of (5.0: 5.0) is usually the fastest pattern of gait practicable to a hexapod; a further reduction of the relative duration of the backstroke, as in figure 4e, results in unstable moments
15 EVOLUTION OF HEXAF'OD LOCOMOTORY MECHANISMS 217 supported by only two legs (figure 40). Gait (5.0 : 5.0) p.d is shown in figure 3a, d, g. A pattern intermediate between this gait and pattern (2.0 : 8.0) on figure 2a with even loading on the legs, is given on figure 3b-c, pattern (4.0 : 6.0) at a p.d. of 0.45 in (b) and 0.50 in (c). All three gaits in figure 3 show reasonably stable time intervals k. The order of footfalls in (d) and (e) are uneven, but differently arranged, and the number of simultaneously propulsive legs varies from three to four and from three to five respectively in the two gaits (a) and (b). Photographic records show that both of these gaits are used by pterygotes, see arrows giving references to the plates. The gait in figure 3c, with a phase difference between successive legs of exactly half a pace duration (0.5), results in the simultaneous change of three supporting legs twice in every pace (f), and the numbers of momentarily propulsive legs changes abruptly from three to six. This gait has not been recorded for hexapods and, if it occurs, is probably not common. This is not surprising, since the mechanical conditions it brings are sufficiently disadvantageous to preclude its use for anything beyond momentary periods of time, but this gait is frequently used by the arachnids (Part 11). The lines marked X-Y on figures 2 and 3 pass through successive changes of supporting legs in the given gaits. When p.d. is 0.50 this line is vertical (figure 3c). When the p.d. is <0.5 the lower end of the line slopes to the right (figures 2a-c and 3b) but when the p.d. is >0.5 the slope is in the opposite direction (figures 3a, 5a). The line X-Y also indicates the direction of the shift of the metachronal wave over the body; whether it goes from tail to head, as is usual, or from head to tail, as is less common, has no propulsive significance.* The possibility of a vertical line X-Y does not arise except in the fastest stable patterns of gait, the (5.0 : 5.0) and the unstable (5.5 : 4.5), and is then avoided by increase of the p.d. to just over 0.5, as in figure 3a and pl. 1, figure f. Cinematograph records of earwigs running at their fastest show an almost, but not quite, vertical line X-Y, as does the record of fast running in the cockroach by Hughes (1952, text-fig. 7 lower diagram), both of which have an approximate gait pattern of (5.0 : 5.0). Figure 3f shows two groups of simultaneous footfalls per pace, L.l, L.3, R.2 and L.2, R.l, R.3 at a phase difference between successive legs of As this phase difference is reduced from 0.47 to 0.2 in figure 4d-a, the footfalls of legs 2 and 3 occur at progressively later time intervals after the footfalls of leg 1 (figure 4i-f), so that their positions along the pace lines shift to the right in gaits (d) to (b). Uneven footfalls intervals are present in gait (d) and (c) and they become even in gait (b) (figure4i, h,g). In figure4f a further shift of the footfalls of legs 2 and 3 has occurred, and in the same direction, so that now the footfalls of legs 3 take place after and not before those of legs 1 of the opposite side. This results in the loss of the bilateral alternation of footfalls L.l, R.2, L.3, R.l, L.2, R.3 seen in gaits (b) and (d) and its replacement by L. 1, R.3, R.2, R. 1, L.3, L.2, an order occurring in very slow patterns of gait where there are only one or two legs off the ground simultaneously, as in figure 4k gait (a) (see also Protura below). A gait intermediate between those in figure 4a and b is one of pattern (2.4 : 7.6) and p.d. 0.3, which shows footfalls The transmission of the metachronal wave in Onychophora is marked by the lime r in Part 1, text-fg. 1, the wave travelling forwards in la, b and backwards in lc.
16 218 S. M. MANTON of legs 3 to occur simultaneously with those of legs 1 on the opposite side. This gait has been recorded by Hughes (1952, text-fig. 7 top right, but with different descriptive terminology). When the phase difference between successive legs is >0.5, as in figure 4e (a rare condition in hexapods), the shift of the footfalls of legs 2 and 3 is in the opposite direction, to the left of the footfalls of legs 1, as in figure 4j, and not to the right as in figures 4f-i, thus occurring earlier during the pace than those of legs 1. The number of legs which are simultaneously pushing against the ground is shown by figure 4k-o for the several gaits. Only in the rare gait (e), with a relative duration of backstroke of less than half a pace duration (5.5 : 4.5), are there unstable moments with only two legs in contact with the ground (figure 40). Such moments are short and the pace duration at these fastest gaits is also short, so that there is enough momentum to carry the insect over the two moments of instability per pace. It will be noted that any hexapod gait showing a time interval k (marked on figures 2, 3, etc.) will at the same moment show two successive legs in the propulsive phase, converging towards one another, as in epimorphic Chilopoda (Part 3, text-fig. 3, pls. 32, 33). This occurs whether the p.d. is >or<o.s, the converging propulsive iegs having their footfalls on the same groups,of footprints (figure Sa, b, g, i). At other moments during the pace, when the gait pattern is slow enough to show two successive legs in the propulsive phase, these legs diverge, their footfalls being on successive and not on the same group of footprints (figures 5, 6, 7). 4C. CONCLUSIONS CONCERNING THE GENERAL CHARACTER OF HEXAPOD GAITS Thus, to the list of well known requirements for hexapod stability in running, namely:- a minimum of three supporting legs whose tips are arranged in a triangle; the paired legs moving in opposite phase; and legs 2 and 3 being put on the ground either before or simultaneously with the raising of legs 1 and 2 respectively on the same side; we can add the following:- the most mechanically advantageous hexapod gaits giving least strain on the organism are those in which the footfalls occur at equal intervals of time, and which thereby produce even loading on the legs. The choice of gaits by the Chilopoda, and the number of legs used by the adult and by their larval instars, is determined by these same factors (Part 3). But in the hexapods these advantages accrue only during the slower patterns of gait, and must be given up in some measure if greater speeds of running are to be obtained by changes in the pattern of the gait. Harmonious changes in the phase differences between successive legs accompanying changes in the patterns of the gaits are obligatory maintaining suitable periods of time during which two successive legs are in contact with the ground (marked k on figures 2b, and 3a-c). The actual order of footfalls upon the ground and the leg loading is determined by the pattern of the gait and the phase difference between successive le s. All recorded gaits of hexapods, exclu dg ing the jumping gaits of the Thysanura, form parts of the described series of gaits which exist for mechanical reasons, and no one species has been found to employ gaits with striking individual characteristics which do not fall within these obligate series.
17 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 219 PTERYGOTA Relative durations of forward and backstrokes 2.0: 8.0 Phase difference between successive legs a I PIlh b Figure 2. The hexapod gait diagrams shown here and on other figures, share the same conventions. The numerical entries for the relative durations of the forward and backward strokes are given as in previous Parts, 10.0 representing the duration of one pace. The entries for the phase difference between successive legs represent that proportion of a whole pace by which leg n + 1 is in advance of leg n, 1.O here representing the duration of pace. The heavy lines depict left legs during their propulsive phase, in contact with the ground, and the thin lines show the forward swing. The movements of the right legs are shown by interrupted lines. All movements shown are relative to the head of the animal and progress is from left to right. The line X-Y passes through successive changes in leg support, here by left legs 1 and 2 followed by right legs 2 and 3, for further description see text. (a-c) Gait diagrams to show the execution of the same pattern of gait at three alternative phase differences between successive legs. The speed of walking in all three gaits is similar if the pace duration is the same. On (b) a horizontal line k marks the time during which both left legs 1 and 2 are simultaneously on the ground and occupying the same group of footprints. (d-k) The horizontal lines in each represent the duration of one pace; (d-i) lie below and between successive footfalls of left leg 1, see above (a-c), and (i-k) lie below and between successive footfalls of right leg 2, see above in (c). The footfalls of the three right and left legs are marked on either side of the horizontal pace line in (d-f) and in (g-i) are marked the number of legs which are simultaneously propulsive in the given time intervals, six legs being in use. In the Protura, walking on legs 2 and 3 only, the order of footfalls is shown in (j) and the number of legs which are simultaneously propulsive in (k). The arrowed lines below (c) show the moments in time of the photos of F'rotura in pl. 1, figures i, j, k, legs 2 and 3 only being used.
18 220 S. M. MANTON PTERYGOTA TlMt Relative durations of forward and backstrokes 50: : :6.0 Phase difference between successive legs C Figure 3. Conventions are as described for figure 2. A range of hexapod gait diagrams (a-c) illustrating the relationship between the relative durations of the forward and back strokes and the phase differences between successive legs. (a) Shows a pattern of gait (5.0 : 5.0), which is usually the most speedy pattern practicable to a hexapod. A slower pattern (4.0 : 6.0) is shown in (b) and (c) at phase differences between successive legs of 0.45 and 0.50 respectively. (d-i) Gives the resultant order of footfalls and numbers of propulsive legs at successive moments throughout a pace. The latter in (e, f, h, i) are very different for gaits (b) and (c) although the pattern of the gait is the same. Photographs of a beetle, a bug and an earwig performing gaits (a) and (b) are shown in pl. 1, figures a, c, f the moments being marked by arrows below (a) and (b). For further description see text. No examples of hwapods using gait (c) have been recorded, but arachnids frequently use this gait, see Part 11.
19 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 221 PTERYGOTA SLOWER PATTERNS OF GAll > - < FASTER PATTERNS OF GAIT LEFT I Relative durations of forward and backstrokes I I I 1.5:8.5 I 2.5:7.5 I 3,516.5 I 4.5: 5.5 I 5.5:4.5 I I I I Phase difference between successive legs I 0.2 I I I I I! 0.6 RIGHT FOOTFALLS f TIME I I I I I 1I I i 3 1 I h I i I I I I 5 p;.5 fi 5 +,5 5,+ 514,5,4 p,4 $14 I5 I4 pi I,4,&4,514 3,41wr(l I I 3,Zl~Wl?r 3 mp12 I1 I1 11 I II PDCS NUMBERS OF PI(OPULSI*E LEGS I I I I k I rn n 0 I I I I i Figure 4. A range of gaits from fast to slow occumng among pterygote insects are illustrated by the data assembled under the gait stills (a-e). The relative durations of the forward and backward strokes and the phase differences between successive legs for each gait are given below the upper gait stills. These depict the disposition of the legs for the moment in each gait when the right leg 3 is at the end of the backstroke. The heavy lines indicate legs in the propulsive phase and the thin lines show legs performing the forward recovery swing off the ground. For convenience the legs are drawn at equal lengths and swinging about the same angle on the body, the range of movement of the tip being shown by the heavy lines on the right of (e). Neither of these features exactly obtain, the legs being of unequal lengths although taking the same stride (see Part 2, figure 3 Rorficuh). From a fuller series of such diagrams an approximate estimate of a gait can be made from a limited number of still photographs, (see 5 2). The conventions in (f-o) are similar to those described for figure 2 and give data for each of the 5 gaits shown above. The actual pace duration in the slower patterns of gait is greater than in the faster. The phase differences between successive legs are those that will give a reasonable, but not excessively long or short time interval k, marked on figures 2 and 3a-c, such as is employed by the Pterygota.
20 222 S. M. MANTON 5. THE RUNNING ACHIEVEMENTS OF THE PTERYGOTA Adult Pterygota, when running on the ground are, on the whole, less swift than Myriapoda of comparable size. Where speed is appropriate, the adult pterygote takes to its wings. At room temperature an 11 mm earwig, Forficula auricuzaria L., can achieve 100mm/sec. in contrast to the somewhat larger 15 mm Lithobius forficatus (L.) and 22 mm Scutigera coleoptrata (L.) which reach 280 and 420mm/sec. respectively. Exceptional fleetness is found in Periplaneta americana which at 22OC can run at 660mm/sec. (McConnell & Richards, 1955), but this is a larger animal than the centipedes noted and would be expected to run more speedily; a small cockroach, such as Blatella germanica achieves 293 mm/sec. (loc. cit.). The tiger beetle also appears to be very fleet. The rate of progression when running depends upon the length of the stride and the rapidity of stepping. The former is controlled by the pattern of the gait, the length of the legs, their angles of swing, and the latter by the physiological capabilities of the animal. P1. 1, figures a, b show tracks left by an earwig running over smoked paper, the stride lengths (the distance between the groups of footprints on one side) and the speed being greater in (b) than in (a). Relatively shorter strides are shown by the more slowly running shield bug and the beetle in pl. 1, figures c, d. Each plantigrade tarsus makes a number of marks on the paper and often a crescentic sweep forwards as the leg is raised, so that the tracks are less easy to read than those of unguligrade apterygotes. The rapidity of stepping can vary considerably without any change being made in the gait. The pace durations of Gastrodes ferrugineus. for example, can be doubled, so reducing the speed to half, without any other changes. Hughes (1952) noted that Blatta does the same sort of thing. This contrasts with the Onychophora, Chilopoda, etc. where changes in the rate of stepping are usually accompanied by changes in the gait. It has been shown above that only medium to slow patterns of gait are practicable to most hexapods, the pattern seldom being faster than (5.0 : 5.0). Part 2, Table 1, p. 104 shows a comparison between the speeds of the earwig with those of certain Chilopoda, Diplopoda, a spider and Peripatopsis, giving also the contribution towards increase in speed made by changes in the pattern of the gait, by the pace duration and by the angle of swing of the leg. At the slower speeds the earwig increases its speed of running mainly by shortening the pace duration (stepping more quickly) and at the faster speeds the changes in pattern of the gait become more important, but increase in the angle of swing of the leg (stepping out) provides the greatest factor towards fastest running. Actual ranges of gait pattern recorded by cinematography (needing bright lights) were: for Forficula auricularia (4.5 : 5.5) to (5.0 : 5.0); for the hemipteran Gastrodes ferrugineus (1.75 : 8.25) to (4.75 : 5.25); and for the beetle Cylindronotus laevioctostriatus (2.5 : 7.5) to (4.75 : 5.25). In dim illumination most insects use slow patterns of gait. The pace durations of Forficula under bright light ranged from 0.2 to 0.08 sec.; for Gastrodes 0.23 to 0.07 sec. over a speed range of 15 to 57 mm/sec.; and for the beetle 1.2 to 0.4 sec. over a speed range of 2.4 to 12.8 mm/sec. The pace durations used by smaller insects, such as ants, are often much shorter than those of the larger species.
21 EVOLUTION OF HEX24POD LOCOMOTORY MECHANISMS 223 Some Pterygota habitually, or at times, walk on only two pairs of legs, Hughes (1952) noted the praying mantis walking on legs 2 and 3, while the hopper stages of some locusts walk on legs 1 and 2. The gaits employed are slow in pattern, a gait such as that on figure 3a would be impossible, because of long periods of support by only two legs. Butterflies walking about on composite flowers often do so slowly on four legs, one at a time being raised (cf. Protura below). A greater divergence from the usual pterygote gaits is found in certain larvae, such as those of staphylinid beetles. The larvae live in soil, and therefore need to have a minimal leg projection laterally or leg movements would be impeded by the soil, as for myriapods (Parts 3 and 4). Locomotion on the surface of the ground is assisted by a punting action of a median peg at the end of the staphylinid abdomen, resulting in long strides being taken by the short legs, shown well by the track on pl. 1, figure g compared with the shorter strides taken by the shield bug and earwig, (a) and (c), both possessing relatively longer legs. When abdominal punting is most active in the staphylinid, the paired legs move in approximately the same phase (footprint groups 4 and 5 on pl. 1, figure g are almost level), but the transverse coordination is very flexible and footprint groups 1-3 on this track are roughly alternate. When the abdominal punting is absent (pl. 1, figure h), then the paired legs move in opposite phase and the stride length is less than half the length shown on the right of pl. 1, figure g (the moment is marked on figure 3a by the arrowed vertical line). 6. LOCOMOTORY ACHIEVEMENTS AND HABITS OF THE APTERYGOTE HEXAPODS In an ability to run very speedily most of the wingless hexapods, other than Thysanura, have at the present day been outstripped by the fleetest of the carnivorous myriapods. It is probable that the initial differentiation of a hexapod, with the advantage of longer legs facilitating faster running, came into existence long before the extreme present-day specializations of the myriapods. The usual limitation of hexapods gaits (except for Thysanura) to the slower patterns, giving stability at all moments in time, necessitates in the apterygotes a use of rapid stepping, or of wide angles of swing of the legs, or other means of obtaining agility sufficient for escape from more rapidly moving predators, such as winged insects, arachnids, birds, etc. A parallel evolution of an escape jumping habit, by different means, has been acquired by Collembola and by Thysanura. Efficient protection is gained by the ability to jump repeatedly in any direction, and many times the height of either predator or prey, so making it difficult for a predator to follow visually. A second response to the pressure set up by other animals appears to have been that of secondarily acquired deeper-living in the soil, decaying logs, etc. where long legs are unsuitable. Plenty of short-legged Collembola can no longer jump, their jumping organs and associated muscles being in various stages of regression; japygids are suited to crevice life and not to surface locomotion (9 6 A(i)); Lepismatidae are unique in their achievement of very rapid and wide movements by the distal part of shortish legs, so providing speedy running in both open and restricted spaces ( 8 6 C(v), 7 C(iii)).
22 224 S. M. MANTON 6A. DIPLURA Representing this class, species of Campodeidae were collected round London, and live japygid species from the Solomon Islands were sent to me by Mr and Mrs Greenslade, working on the British Museum (Nat. Hist.) expedition to this region. (i) Habits and habitats of the Diplura These hexapods live under cover during daylight. There is no doubt that the Campodeidae live predominantly in highly porous soils, and that they negotiate existing crevices by their small size and flexibility of body and not by pushing. The Japygidae live in deep humus beds, in ground litter and under stones. They do not press against the soil with their bodies, but burrow by using existing crevices, great flexibility of trunk and very strong leg movements. They are most competent scrabblers. In the Solomon Island soil samples the japygids were usually found inside lumps or crumbs of soil about 5/2 inch in diameter. When placed on a soil surface the animals at once go down a pore or crevice and can be watched through a glass vessel as they most readily twist and turn in small spaces. The Campodeidae are remarkably fleet (see below) and they use this ability at least to seek out local damp situations when the general environment is desiccating (J. G. Blower, pers. comm.). The japygids are not fleet but are highly adapted towards their proficiencies in the soil. Marten (1939) records collecting Campodea lankesteri Silv. and C. colladoi Silv. in Germany below large heaps of decaying leaves on the stable surface between leaves and earth. Van der Drift (1951) gives the following figures for numbers of Campodea per m2: in the top 1-8 cm of new and old litter, 1.5 Campodea; below in the 1-4 cm of amorphous black material with few recognisable leaf fragments, 59.1 Campodea; and in the underlying 1 cm of mineral soil with some humus, Campodea. In Blower s monthly sampling from a British Sycamore/Ash wood, with open canopy and patches of soft grass (Holcus) on the ground, he recorded 12.5 times as many Campodea plusiochaeta per m2 in soil free from recognisable organic debris as in the litter, or grass roots, or other surface debris (pers. comm.). In these temperate environments the campodeids are mainly soil inhabitants. On the contrary, in the tropical rain forest of the Solomon Islands, about four times as many campodeids were recovered from the litter than from the soil below, while the japygids were more abundant in the soil than in the litter (Greenslade, pers. comm.). In these forests about 2 inches of litter lies above a?h inch of powdery humus over the mineral soil. In tropical rain forest organic matter is concentrated close to the surface and the mineral soil below is fine textured with little crumb or pore formation, and is likely to be less easily penetrated by small animals than in a temperate woodland where the mixing of mineral and organic material goes to a greater depth, so that the corresponding inhabitants of the tropical upper layers may occur deeper down in temperate forests. (Greenslade, pers. comm.). The penetration of soil spaces is facilitated by the extreme flexibility of the thorax, both dorso-ventrally and laterally, particularly in the japygids. Here the prothorax can remain in line with the head, as in pl. 2, figures f-h, but the two
23 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS can flex horizontally on each other so that an angle of 50 lies between them. Even greater freedom lies at the pro-mesothoracic junction where the one segment may turn on the other by up to 90, this being the most flexible intersegment in the body (pl. 2, figures el i). The head can swing laterally, then posteriorly and cross over the metathoracic tergite to face towards the other side without movement of the standing metathoracic legs. This thoracic flexibility is assisted by the small diameter of the intersegments and by the skeleto-musculature described below ( 5 5 7A, 8B). The narrowness of the thoracic segments compared with the head and abdomen, particularly in the japygids, enables the legs to work close in to the flanks without lateral projection. The legs of japygids are not longer, relative to thoracic segment volume, than in many epimorphic Chilopoda where such good shelter cannot be obtained. When walking on the flat the legs project freely (pl. 2, figure h); The Diplura are carnivorous, scavenging at times, and have also been recorded consuming decaying vegetable matter and fungal mycelia (Denis, 1949). Campodea lankesteri shows preference for the larvae of the midge Sciara but eats other animal food in the soil (Marten, 1939). In central Europe the japygids show a preference for soft sluggish onychiurid Collembola, but also take campodeids. The flexibility of thorax and abdomen is used in feeding, prey being caught either by mouth parts or by the terminal forceps, a bending of the body bringing the food and forceps to the mouth parts (Kuhnelt, 1961). The terminal cerci in campodeids serve as posterior antennae, just as do the 15th pair of legs in Scutigera. 225 (ii) Telescopic antenna1 movements of the Japygidae One of the most striking accomplishments of japygids is the actual shortening and extension of the antennae as they feel about in the soil. The differences in the lengths of antennae in pl. 2, figures d-g are real and not due to foreshortening. An extended antenna can shorten to 2/3rds of its length by the telescoping of each segment into the next. Such an ability has not been found in another hexapod. The intrinsic antennal musculature of a japygid was first described by Imms (1939) from material which I collected in South Africa, but on which no observations were made in the living state. He contrasted this dipluran condition with that of the Pterygota, where all the antennal musculature is extrinsic and no intrinsic muscular movements exist. He figures dorsal and ventral retractor muscles passing between one antennal segment and the next, also some more oblique extensor s muscles and more elaborate sets of muscles in the basal segments. These muscles doubtless account for the antennal shortening shown in pl. 2. But the extension of an antenna is more likely to be effected by blood pressure than by the inadequate complement of muscles claimed by Imms to be extensors. Imms (1939) described and figured an enormous sinus-like blood vessel lying in the centre of the antenna and constricted intersegmentally. The inflation of each segmental chamber along this vessel, under the control of its own muscles, could provide a mechanism for antennal extension of great flexibility, in that the segmental chambers along the vessel could dilate independently, so changing the shape of the antenna. Such antennal blood vessels have not been
24 226 S. M. MANTON recorded elsewhere, suggesting that their presence may be correlated with telescopic movements, serving to extend the antenna, either locally or as a whole, while muscles are principally responsible for shortening and for flexing the antenna. The anatomy and movements of the japygid antenna represent a high degree of specialization used in exploring minute crevices. The antenna of Gzmpodea is also muscular within, but no telescoping has been recorded, although the antennae are very flexible. (iii) The running achievements of the Diplura The locomotory abilities of the Campodeidae and Japygidae are very different, although the gaits employed are much the same. The effects of the differences in leg length in the two groups has been shown in figure 1 and 5 4A, the long-legged Campodeidae being decidedly swift, while the Japygidae are not. In Gzmpodea the range in pattern of the gaits in a bright light, as used for cinematography, was from (5.5 : 4.5) to (2.5 : 7.5), the phase difference between successive legs changing from 0.6 to 0.33 (figure 5a, b and pl. 2, figures a-c). The transition in support by successive legs on the same side is not quite simultaneous, but the time interval k, marked on the figure 5b, is brief, cf. figures 2 and 3. Much slower patterns of gait are probably used in darkness in the soil where spaces are confined. This fairly wide range of gaits (see Part 2, p. 105) facilitates varying the speed in running. The fast locomotion upon exposed surfaces depends on the long strides taken by the long legs (figure 1 and 5 4A), facilitated by the great differences in leg length of the three pairs, so allowing the fanning out of the fields of movement shown in figure 5j. Unstable gaits are rare in hexapods, and in the fastest pattern used by Gzmpodea (5.5 : 4.5), there are brief moments when the body is supported by only two legs (figure 5e), but the unstable moments are of short duration. The animal appears to extract the maximum speed allowable by its morphology. The speeds achieved under bright illumination by a Gzmpodea of body length 4.5 mm ranged from 18 to 54 mm/sec., a minimal pace duration being 0.05 sec. and a maximal stride length 2.7 mm. Gzmpodea is thus agile compared with a 4 mm Lithobius and a 5 mm Scutigerella achieving up to 26 and 42 mm/sec. respectively. The pace duration is not so brief as in the Symphyla and the chilopod Cryptops, but it is shorter than in most Myriapoda and Pterygota. Japygids do not excel at surface running, apparently disliking the exercise. They would not maintain steady walking movements for long enough to record reasonably accurate figures for speeds. Their agile movements are reserved for manoeuvring in soil crevices. During their intermittent walking on flat surfaces they employ much the same gaits as are seen in Chmpodea (PI. 2, figures d-g), but the stride length is relatively shorter, owing to the length of the legs (figure l), and the momentary maximum speeds are much less than those of Gzmpodeu. The gaits employed by Jap~x are noted in the legend to pl. 2. (iv) Conclusions concerning dipluran locomotory achievements The data disclose a remarkable difference between campodeid and japygid Diplura. The latter possess extreme specialization and ability towards
25 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 227 CAMPODEA Relative durations of forward and backstrokes 5.5 : :7.5 Phase difference between successive legs C I I Figure 5. The gaits of Campodea. Conventions are as described for figure 2. Gait diagrams (a) and (b) show the fastest and the slowest gaits utilized during cinematography. (c) and (d) show the sequence of footfalls of the six legs in the gaits shown above. (e) and (0 record the number of propulsive legs at successive moments during one pace of the gaits shown above. (g-i) Gait stills show diagrammatically the disposition of the legs at the moments shown in PI. 2, figures a-c. The legs are drawn as if they were of equal lengths and swinging forward and backward about their bases to equal extents. The extreme forward and backward positions of the legs relative to the base are indicated by the ends of the vertical lines alongside. Actually there is an overlap in the fields of movement, as shown in figure (j). (g) gives the moment in time, marked by the arrow below (a), seen in the photograph pl. 2, figure a, (h) shows the moment in time seen in the photograph pl. 2, figure b and (i) gives the moment in time marked by the arrow below (b), seen in the photograph pl. 2, figure c. (j) The range of movement of the right legs drawn relative to the body, the heavy vertical lines depicting the propulsive backstroke, as in Part 2, text-figs. 2 and 3.

26 228 S. M. MANTON penetration of soil and negotiation of crevices, but not to walking on the flat. Japygids possess a more highly adapted antenna suiting exploration of crevices than occurs in any other hexapod, where telescopic antennae are unknown. The campodeids predominantly live deep in the soil, at any rate during the day time, yet they have long projecting legs such as are not usual in animals which penetrate -deeply, and they display very considerable agility and speed of running by long strides on flat surfaces. The question is thus posed: why do the campodeids need such agility while the japygids do not? Both groups hunt living prey. Possibly the campodeid agility, whereby they can find adequate moisture conditions with rapidity, contributes to their remarkably wide and successful distribution in contrast to the japygids, which, in spite of their great proficiencies, are much more limited in distribution. 6B. COLLEMBOLA The Collembola are the oldest of the known hexapods, fossil remains of Rhyniella praecursor Hirst & Maulik from the Devonian have been described most fully by Scourfield (1940) etc. But, as noted by Jeanne1 (in Grasse, 1949), this animal is already a specialised collembolan. The legs and antennae are short and there is no separate prothoracic dorsal sclerite. (i) Habits and habitats of the Collembola There is a wider range of habitats and habits among the Collembola than in the Diplura. The habits of Collembola, and the environments they frequent, have been summarised by Kuhnelt (1961) as follows. The three most important locations are: (1) The soil surface and porous superficial layer, in litter and moss, under logs etc. Here the species are large, 3-4 mm; ocelli and good springing organs are present and the legs are long, e.g. Tomocerus and Orchesella. (2) The upper soil layers where the species are smaller, 2 mm, heavier in build, with a shortening of the springing organ and the legs, e.g. Folsomia. (3) The deeper soil layers, where legs are short, springing organs absent, ocelli absent and body worm-like and white. Small size enables easy passage through minute cracks and spaces, but no active burrowing is possible, e.g. Tullbergia, Friesea. Greater resistance to desiccation is shown by the species living at upper levels, but aii require humidity, and even Tomocerus can go down to 15 cm in the soil during dry conditions. The food is very various, ranging from decayed plant material, fungal mycelia and spores, animal food such as fly pupae, fly larvae, dead flies, other Collembola, etc. and each species has its own particular preferences which determine to some extent the microhabitat and locomotory activities. (4) Above the soil live the Symphypleona, with globular bodies, small thorax, long legs and long antennae (pl. 1, figure n). They are found on damp wood, Alacma fusca climbs trees at night in Britain and feeds on epiphytes and Sminthunrs viridis can be a pest of crops in Australia (J. G. Blower, pers. comm.). For the present investigation the large jumping Tomocerus longicornis Muller and Orchesella villosa Geof. and the small white Onychiurus sinensis Stach. have been used for the examination of internal and external anatomy and movements. Living Anurida maritima (GuQin), Neanura muscorum Templeton,
 The running abilities of Collembola Both running and jumping are well displayed by the larger and longer-legged Collembola living on the soil surface and in the upper porous layers.")
27 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 229 unidentified Hypogastrurida and small species frequenting the inside of decaying logs have been studied, together with a sminthurid representing the S ymphypleona. (ii) The running abilities of Collembola Both running and jumping are well displayed by the larger and longer-legged Collembola living on the soil surface and in the upper porous layers. In the deeper niches both accomplishments wane, locomotory movements become slower and jumping absent. Precision stepping takes place in Collembola in spite of the hydrostatic mechanism used in jumping. Orchesella villosa Geof. and Tomocerus longicornis Muller under bright illumination almost always used their same fast gait (4.5 : 5.5) at a phase difference between successive legs of about 0.44 (see figure 6a and pl. 3, figures a, el f). But much slower patterns of gait are employed when the animals are pottering about in dark or dimly-lit conditions. Two of these patterns are shown in figure 6b, c, bringing with them the usual advantages of more even loading on the legs, figure 6e, f, h, i. The change in support by successive legs on the same side of the body (ringed points on figure 6a) is almost simultaneous in all recorded gaits, the stable moments marked k on figures 2, 3, 5 being absent. But the ringed points for left legs 1-2 and right legs 2-3 are staggered, conferring stability; they are not exactly above each other on figure 6a. Only slow patterns of gait have 'been recorded from the long legged but small sminthurid (pl. 1, figure n, cf. Tomocerus pl. 3, figure a) and from Anurida maritima, with legs which are shorter than those of Tomocerus but not so short as in many crevice-living Collembola. A commonly used gait of Anurida is shown in figure 6c, the arrow below indicating the moment in time shown by pl. 3, figure g. The sminthurid in pl. 1, figure n is using a slighly slower gait, (3.0 : 7.0) approx. The gait stills in figure 6j, k, 1 show movements in time during the three gaits, (a, b, c), when right leg 3 is at the end of the backstroke and the disposition of the remaining legs is an indication of the pattern and characteristics of the gait ( 8 2, 4B, figure 4). Figure 6j shows a mirror image of a moment later than pl. 3, figure e and figure 6,l exactly correspond with pl. 3, figure g. The speeds achieved under bright illumination by an Orchesella villosa 4 mm long, were <lomm/sec., but the animal can probably do as much as 15 mm/sec. The strides were c2 mm and the pace duration X.2 sec. Tomocerus longicornis, 5 mm long, does a little better with longer legs (see pl. 3, figures a, e), the strides recorded were <3 mm, the pace durations shorter, >0.17 sec. and recorded speeds on the short runs the animal makes were <20 mm/sec. but it can certainly achieve 25 and possibly 30 mm/sec. Collembola with shorter legs run more slowly and the pace duration is often long, as in soil-, crevice- and cave-living hypogastrurids and Neanuru spp. (pls. 3, figures h, i; 5, figures e, 0. The stride is short because the legs are short, pl. 5, figure e. Fast running is not necessary for these animals, and they have greater use for flexibility of body than for an ability to jump. Instead of a hydrostatically rigid body (pl. 3, figures a-f and see below) much twisting and turning is possible, as shown in pl. 3, figures h, i: all useful accomplishments in
28 230 S. M. MANTON TOMOCERUS Relative durations of forward and backstrokes 4, : j Phase difference between successive legs h I Figure6. The gaits of Collembola (Tomocerus) and of some Thysanura (Lepisma and Thennobiu). Conventions are as described for figure 2. The changes in support by successive legs 1 and 2 and by legs 2 and 3 are ringed. Gait diagrams in (a) show the fastest and (b-c) the slower patterns of gait shown by Tomoceiws longicornis Muller. Gait (a) is seen in pl. 3, figures a, e, f the moments being marked by arrows below (a). (d-f) Shows the sequence of footfalls of the six legs in the gaits illustrated above. (g-i) Records the number of propulsive legs at successive moments during one pace of the gaits shown above. (j-i) Gait stills of the disposition of the legs at the moments indicated by the long arrows pointing towards the three gaits shown above. In all stills right leg 3 is at the end of the backstroke, heavy lines indicate a leg in the propulsive phase and thin lines show the forward swing, the legs are drawn of equal lengths and swinging similarly about their bases, the range being marked alongside on the right. The actual ranges are seen in (m). (m) The range of movement of the right legs of Tomocerus Iongicornis'drawn relative to the body, the heavy vertical lines depicting the limb tip during the propulsive backstroke. The small labelled arrows below (a, c) show the moments in time seen on the plates for Tomocerus!and Lipura pi. 3, figures a, e, f, g.
 The collembolan jump When Collembola, equipped with well formed jumping organs, are disturbed, they jump readily and repeatedly, but tire easily, as with jumping Thysanura.")
29 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 23 1 negotiating narrow crevices without pushing. The anatomical basis of different degrees of trunk mobility is considered in and 8. (iii) The collembolan jump When Collembola, equipped with well formed jumping organs, are disturbed, they jump readily and repeatedly, but tire easily, as with jumping Thysanura. The successive jumps take the animals in any direction, cm upward and landing a little distance away. The usual disruptive or cryptic colorations make the jumping movements difficult: to follow by the eye, and the exercise provides protection against predatory birds, etc., disturbing the litter. The mechanism and anatomical basis of the jump are described in 5 9A. (iv) Conclusions concerning the collembolan running achievements The running abilities of the Collembola, although much better than those of the Japygidae, contrast with the much more agile Campodeidae. The Collembola are the only class of aprerygotes lacking a firm articulation, or close union, between a body sclerite and the coxal rim (3 7B(i)). Thus an inability to step quickly might be expected of this group. When speed of movement is required the more surface living species jump. It is shown below how the whole anatomy of body and legs of Collembola is adapted in great detail to provide the jumping mechanism, and the evolution of the trunk within this class is centred on this habit (9 10F, G). The body of the jumpers has need to be rigid and inflexible (9 9A). A secondary loss of the jumping habit has led to regression of the jumping organ, reduction in leg length and an increase in trunk flexibility, all contributing to easy living in crevices ( 5 9B). 6C. THYSANURA Three species representing this class have been investigated: Petrobius brevistylis Carpenter, from the M. B.A. Laboratory, Plymouth and Port Erin, Isle of Man, Lepisma saccharina L. from my house and Thermobia domestica Pack, from the Pest Infestation Laboratory, Slough. The Thysanura comprise hexapods of the largest size among the apterygote classes, and excel both in running and jumping. They possess good compound eyes in contrast to other apterygotes. (i) Habits and habitats of the Thysanura The Machilidae, or rock springers, inhabit rocky shores, dry forests, etc., sheltering between and under stones or leaves. They are not soil animals and they feed largely on Pleurococcus, or other unicellular algae, or by nibbling dry or fresh vegetable matter (Handschin, 1927; Kiihnelt, 1961). The Lepismatidae are regular inhabitants of warm sandy soils (Egypt), they live in association with ants (Europe), as well as in human habitations and food stores. Their food can be dry and hard, such as the grains of cereals, and their jaws are suited to such feeding (Manton, 1964).
30 232 S. M. MANTON Thysanuran locomotion serves for moving to fresh feeding places or to shelter and for escape from predators, but not for hunting live prey. Perfection in running and jumping is on different lines from the locomotory achievements of other classes of hexapods. Both Machilidae and Lepismatidae have exploited the use of unstable jumping gaits, with less than three legs in contact with the ground at one moment. Gzmpodea at times uses unstable gaits (0 6A(iii)) and certain Pterygota perform high jumps, but with one pair of legs only (Locusta). Machilidae normally progress by a series of short jumps or hops, all six legs being used. The tracks left by such jumping are regular (figures le, 8g, h; pl. 5, figures a, b). Machilids can also make high jumps by the use of the abdomen (9 9C) landing some distance away and facing in any direction. These jumps are intermittent and form no regular track. The high jumps appear to be an escape reaction, as are the jumps of Collembola. When machilids run on the underside of rocks, the jumping gaits become inappropriate and are abandoned, normal, stable hexapod gaits then being used. The Lepismatidae achieve great fleetness from not very long legs by moving the distal part of the leg very rapidly through a wide angle of swing in a manner unique to this group of hexapods. Fleetness is combined with very little leg projection beyond the flanks, so facilitating moving in confined spaces. The basic action of each leg is probably the same as in the jumping or hopping movements found in the Machilidae (see below and 9 7C(iii)). (ii) The walking gaits of Petrobius These gaits are used when the animal cannot progress by the jumpinggaits, which do not provide sufficient adherence to the under side of rocks, etc. The (4.0 : 6.0) to (2.5 : 7.5) patterns have been recorded photographically from animals walking on a ceiling of filter paper (figure 7b, c). Change in support by successive tarsi on the same side are not simultaneous, the period k being marked, and there are usually 4-5 legs holding on to the ceiling by the tarsal claws at any one moment (figure 7f, g). The paired legs are in opposite phase, as is usual for hexapodous running, as is seen in pl. 6, figures e, f, g. These photographs also show plainly which legs are in contact with the ground, these limb tips and their shadows meeting, see particularly pl. 6, figure f. The moments in time shown by these photographs are illustrated diagrammatically by gait stills in figure 7h, i, j (see the arrows). The execution of gaits on the under side of rocks, etc., is not so accurately or so speedily carried out asare gaits performed on an upper surface. (iii) The jumping gaits of Petrobius These gaits contrast with the walking gaits of Petrobius, and those of all other hexapods when walking or running as opposed to swimming, in that each pair of legs is used in the same phase (pls. 5, figure a; 1, figures 1, m) and the leg action is one of jumping and not of walking or running (see below). The groups of footprints formed by legs on the two sides of Petrobius lie at the same transverse levels (pl. 5, figures a, b) and are not staggered alternately as in other hexapods (figure 1; pl. 5, figures c, d, e, etc. and pl. 1, figures a, b, c). A plan of part of the track on pl. 5, figure a is drawn on figure 8g.
31 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 233 WALKING GAIT Relative durations of forward and backstrokes 4, : 75 Phase difference between successive legs hi I I Figure 7. Petrobius brevistylis Carpenter, walking gaits. (a) Shows the range of movement of the legs relative to the body, the heavy lines depict the legs tips during the backstroke when they are in contact with the ground. During the recovery forward stroke, (not shown) the tips of the limbs are further from the body because each leg is outstretched. Gait diagrams (b-c) record the gaits used when walking upside down on a ceiling of filter paper, conventions as described for figure 2. Both are slow patterns and that of (c) is the slower of the two. Right and left legs of a pair are in opposite phase. The marked arrows show the positions of the moments in time seen in pl. 6, figures e, f, g. These gaits contrast with the normal jumping gaits used on flat or inclined surfaces (figure 8). (d-e) Show the sequence of footfalls of the six legs in the above gaits. (f-g) Record the number of legs in the propulsive phase at successive moments during one pace of the gaits above. (h-i) Gait stills at three moments during the two gait diagrams above, which are seen in pl. 6, figures e, g, f respectively. The legs drawn in heavy black are in the propulsive phase and the diagrammatic range of movement of each leg relative to the body is indicated by the vertical lines on the right of each figure, the swing being equal in the forward and backward direction about the leg base. The actual range of limb tip movement during the backstroke is shown in (a) where the heavy vertical lines are drawn relative to the head.
32 234 S. M. MANTON PETROQIUS JUMPING GAll Relative durations of periods when the tarsi are off and on the ground. 6.7: :5.0-b : '. 0...*.... 2' * h 'L 3 a b Phase difference between successive legs 0: P1.5a PI.11 R1.5~ right ' 3 2 c I i - numberr of d legs e I 5 0' ' I ,213~ ~2 I I I I I I I I I I I I ' 4, 2,4, I J I 6.0:4.0 Figure 8. Petrobius brevistylis Carpenter, jumping gaits. Both legs of a pair are in the same phase, cf. all other hexapods, figures 2-7 and pls (a) Three alternative gaits are shown by a composite gait diagram in which horizontal rectangles indicate the period of time during which a leg is in contact with the ground. The backstroke starts a little before the leg strikes the ground, and the recovery forward stroke, unmarked, lies between successive rectangles. Time progresses from left to right as shown by the horizontal arrow. The relative durations of the periods of time during which each leg is off and on the ground in each gait are given above, the black rectangle being the fastest gait (6.37 : 3.3), the rectangle marked by interrupted lines being the intermediate gait (5.0 : 5.0) and that marked by dotted lines being the slowest gait (3.3 : 6.7). All have a phase difference of 0.33 between successive legs, so that footfalls, marked by large rings, occur at equal intervals of time, see (b). The footfalls of legs 1 are followed by those of legs 3 and finally by those of legs 2. (b) Shows the sequence of footfalls during the three gaits illustrated in (a). (c-e) Shows the numbers of propulsive legs at successive moments throughout a pace in the three gaits. (f) Shows the number of supporting legs in gait (6.0 : 4.0). Slow pattern gaits approaching but not reaching (3.3 : 6.7) have been recorded and the fast (6.7 : 3.3) and the intermediate patterns (6.0 : 4.0) to (5.0 : 5.0) are commonly used. (g-h) The plan (g) of four successive groups of footprints from the track shown in pl. 5, figure a starting from the bottom edge of the white stripe; length of animal 10 mm, stride length 6.4 mm, pattern of gait (5.0 : 5.0) approximately. (h) Plan of four successive groups of footprints from the track shown in pl. 5, figure b scaled to correspond with body length 10 mm, stride 8.3 mm, pattern of the gait between (6.0 : 4.0) and (6.7 : 3.3). The four paces shown on the tracks correspond with the four paces marked by oblique lines on (a). In (g) the groups of footprints are more nearly transverse in position, indicating that legs 2 were put on
33 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 235 As far as has been ascertained, the jumping action of each pair of legs contrasts with the running movements of other hexapods shown by figures 2, 3, 5, 6, 7 in that the backstroke starts before the limb tip touches the ground, as in a galloping horse, so delivering a maximal jumping thrust against the substratum at the moment of contact, and there is a very rapid forward stroke, usually followed by a short rest period. These two features of the jumping movements are seen in pl. 5, figure a (see also 5 7C(iii)). The exposure time of 1/60th sec. gives a blurred image of the most rapidly moving parts, the trunk and legs 2. Only legs 1 are on the ground, delivering a jumping thrust and making a double claw mark, as seen on the right, and hence the claw is stationary and the image of the limb is fairly sharp. Legs 2 give a wide blurred image as they leave their footmarks and swing forwards to their most anterior positions (figure 8a) during the one-fifth or less of the pace duration, covered by the exposure time. Legs 3 are off the ground but are not blurred, therefore they must be going forward more slowly than the body. They have probably just started the backstroke, and their footfalls will take place lateral to those of legs 2, which are visible on the track in front of legs 3, when the forward movement of the body brings legs 3 to this level. These movements cannot be shown in detail by gait diagrams as on figures 2, 3, 5, 6, 7 because the leg movements are too rapid for obtaining precise time periods without high-speed cinematography. P1. 5, figures g, h show two successive frames at 32 frameskc. Only limb tips on the ground are at all sharp at this speed, and the close similarity between the disposition of body and legs in the two frames indicates that a whole cycle of leg movements has occurred in the time interval between the two exposures, legs 3 in (h) being on the ground and one pace in advance of their position in (g). Much more information has been obtained by still photography. Figure 8a shows three alternative patterns of jumping gait for legs 1, 2 and 3. The propulsive jumping thrusts by each pair of legs are delivered against the ground at even time intervals in all gaits, the phase difference between successive legs being The start of a jump by legs 1 is ringed on figure 8a and is followed by legs 3, also ringed, and then by legs 2. The horizontal rectangles mark the period of time during which the legs are in contact with the ground. In fast jumping the backstroke probably starts in advance, to the left, of each rectangle. The gaits are characterized here by the relative durations of the periods when the limbs are off and on the ground: gait (6.7 : 3.3), the fastest, marked in black; gait (5.0 : 5.0), a slower gait, marked by interrupted lines; and gait (3.3 : 6.7), slower still, marked by dots. A phase difference between successive legs of ensures an even rate of progression by the body. the ground very soon after legs 1 were raised, as in gait (5.0 : 5.0), in (h) the groups of footprints are forwardly staggered, indicating that legs 2 were put on the ground long after legs 1 were raised, the travelling body carrying legs 2 forwards off the ground while support was given only by legs 3, as in gait (6.7 : 3.3) shown in (a). The arrows below the gait diagram (a) mark the moments in time seen for Pefrobius in the plates: the arrowed line on the left, passing throu& the interrupted line marking gait (5.0 : 5.0). see pl. 5, figure a; the arrowed line in the middle, passing through the solid line marking gait (6.7 : 3.3), see pl. 1, figure a; and the two arrowed lines L and R on the right mark the moment for the left and right legs of Lepismo, pl. 5, figure c, the paired legs being in opposite phase (not in the same phase as in Pewobius).
34 236 S. M. UANTON In the fast pattern (black) and the slower pattern (interrupted lines) legs n + 1 are put on the ground long after legs n are raised and not before this event, as in most running gaits of other hexapods. Thus the time interval k, conferring stability to the running gaits (figures 2, 3, 5, 6, 7), is absent. In the slowest pattern (dotted lines) legs n remains on the ground until the jump by legs n + 1 starts. The time interval between the footfalls is equal in all gaits (figure 8b) and the numbers of propulsive legs at successive moments throughout each pace are marked in figures 8c, d, e. The fast pattern gait (6.7 : 3.3) is unstable at all moments, only two legs (one pair) at a time being in contact with the ground. Gait (5.0 : 5.0) possesses alternate moments of support by two and four legs, while gait (3.3 : 6.7) is stable at all moments, with two pairs of legs in contact with the ground. The gait commonly used during slower progression is the (5.0 : 5.0) pattern (interrupted lines). The overlap in the propulsive phases of legs 1 and 2 (figure 8a) performing paces 3 and 2 respectively and of legs 3 and 1 performing pace 2 and 3 respectively, and of legs 2 and 3 performing paces 3 and 2 respectively, gives three moments of stability by four legs per pace (figure 8d). The paces on figure 8a are marked by oblique arrows on the left; the three footfalls opposite any pace arrow form one group of footprints on the track. This pattern of gait is seen in pl. 5, figure a, the moment in time is marked by vertical arrows on figure 8a passing through the small circles on the gait marked by interrupted lines. Slower patterns are used, but the extremely slow gait (3.3 : 6.7) shown by dotted lines on figure 8a which is entirely stable (figure 8e), has not been recorded. A commonly used faster pattern than the (5.0 : 5.0) is the (6.0 : 4.0), which is not drawn in full, but is intermediate between the solid rectangles and the interrupted lines on figure 8a, and which possesses short moments of stability with two pairs of legs simultaneously on the ground (figure 8f). The fastest pattern (6.7 : 3.3) or near it, is only used at speed, with short pace durations, doubtless because of its unstable nature, see the vertical arrow through the small circles on the gait marked by solid rectangles in figure 8a and pl. 1, figure 1. Any further reduction in the relative duration of the backward stroke of the legs would result in an airborne moment, and such has not been found in a gait consisting of a regular series of jumps. In the large intermittent escape jumps, which bring the animal high off the ground, the animal is airborne (0 9C). Analysis of the jumping gaits has been made by coordination of all types of evidence, but that of cinematography alone at normal speeds, either in lateral or dorsal view, is very limited in usefulness. Much detail can be obtained from still photographs, using a variety of exposure times, of animals making their tracks on smoked paper. A sufficiency of photos by all methods, gives the range of movement of each leg relative to the body (figure 7a). This, and the legs in sharp image on the photographs, together with the footprints, show exactly at what period in the cycle of movements each pair of legs is in, and gait diagrams show which pattern fits the data. The track in figure 8g from pl. 5, figure a was made by legs 2 being put on the ground soon after legs 1 were raised, etc. (interrupted lines on figure 8a) and the stride or jump length of each leg pair is 6.4mm for the 10mm Petrobius. In figure 8h the stride or jump length is 8.3 mm, legs 2 were carried forward by the moving body for a considerable period after legs 1 were raised
.")
35 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 237 (figure 8a solid rectangles) and the footprint groups are thus forwardly staggered. The maximum recorded stride or jump length is 9 mm for a 10 mm Petrobius and doubtless corresponds with a gait pattern near (6.7 : 3.3). The pattern making the tracks shown in figure 8h is a little slower, between (6.0 : 4.0) and (6.7 : 3.3). The tracks made by the faster patterns of gait, conferring faster speed, are narrower than those made by slower patterns, note the greater width of the track in figure 8g than in (h). This same phenomenon was found in the myriapods, cf. tracks of Callipus running slowly and fast, Part 6, text-fig. 2d and g, where the track is narrower and the stride longer in (d). The placing of the footfalls closer towards the middle line of the track facilitates a wider angle of swing of the leg (as shown in Part 6, text-fig. 3d, cf. the lines df-db and bf-bb explained on p. 498*), but greater effort has to be expended in leg flexure during the backstroke. The fields of movement of legs 1 and 2 are largely anterior to the leg bases while those of legs 3 lie posterior to the leg bases (figure 7a). The thrust against the ground by legs 2 followed by legs 1 (figure 8a) throws the head end of the body upwards and the abdomen downwards (pl. 6, figure b), while the thrust from legs 3 tips the head end downwards and the abdomen upwards (pl. 6, figure c). The pitching movement of the body of Petrobius caused by the jumping gaits (pl. 6, figures b, c), and the general instability due to only one pair of legs being in contact with the ground at a time in many gait patterns, is ameliorated to some extent by the abdominal styli. They have weak extrinsic muscles (see 5 9C) and execute promotor-remotor movements, but they cannot be propulsive organs because they are dragged along the ground leaving interrupted lines on smoked paper (pl. 5, figures a, b). The interruptions presumably correspond with the moments when the abdomen is raised, as in pl. 6, figure c, when most styli are off the ground. The slowing up of the animal as it comes to rest after its last jump by legs 2, and the animal in a prolonged rest period, are shown in pl. 6, figures b-d and (a) respectively. The styli are in various positions, suggestive of a stabilising effect upon the body. The stationary animals in pl. 6, figures I, d hold the abdomen half way between the extreme positions of pitching (seen in b-c) and their large 9th pair of styli are vertical and in contact with the ground, cf. (b). The speeds achieved by the jumping gaits of Petrobius, as determined by cinematography, were from mm/sec. by animals of 10mm body length. Under dim light slower jumping progression occurs. The minimal pace durations were about 0.03 sec. and maximum stride or jump lengths 9 mm. No exact speeds were recorded for the walking gaits practised upside down on a ceiling, but they are not very speedy, either in pattern of gait or in achievement. Conclusions. The walking gaits of Petrobius, used on the underside of objects, closely resemble those of the Pterygota, but the jumping gaits, in which all three pairs of legs are used and each pair is moved in similar phase, are unique among hexapods. These jumping gaits giving speeds of over In Part 6, p. 498 line 9 from the bottom, for smde length read distance travelled during the backstroke.
36 238 S. M. MANTON 200 mm/sec. by a 10 mm animal are indeed rapid when compared with those of the Diplura and Collembola. The fastest running by these animals is that of Campodea, and if such an animal existed with body length 10 mm, it might be expected to run at 120 mm/sec. at maximum. The abdominal styli of Petrobius act as stabilizing organs, assisting the jumping gaits. These outstanding achievements of the Machilidae are dependent upon facilitating morphology of limbs and trunk (see 9 0 7C, 9C). (iv) The high jumping escape reactions of Petrobius When disturbed, Petrobius jumps high off the ground, landing some 3-10 cm away. There is little control of orientation by the animal during the jump, apart from landing upon its feet. On landing, the head end may point in any direction, but the animal may have a sense of direction, because if it lands pointing in the way it came, it sometimes turns about and runs off by the normal gait in the original direction of locomotion. A high jump may be repeated several times, with no running in between, as in Collembola, and in both classes the animals soon tire of jumping. The mechanism of the high jump is considered in 9 9C. (v) The gaits of the Lepismatidae Both Lepisma saccharina and Thermobia domestica employ somewhat the same type of gait as seen in Petrobius, but the paired legs move in opposite phase, and therefore the groups of footprints from the two sides alternate (5 4(i) and figure Id, e). When running their fastest, the groups of footprints of the Lepismatidae are forwardly staggered (pl. 5, figures c, d), a condition found outside the Thysanura only in the tracks of Campodea. When running more slowly the forward staggering is less, see the middle region of the track on pl. 6, figure h, as in Petrobius pl. 5, figure a. When running fast the gaits of the Lepismatidae are performed very exactly, giving the regularity in track seen in pl. 5, figures c, d. But pl. 6, figure h shows irregularity in the degree of forward staggering of the footprints and in their transverse alignment. The lower end of this track records movements in opposite phase on the two sides, in the middle the right and left footprints are almost level, indicating that the paired legs were then moving in almost the same phase, and, at the moment of photography, the alternate movements were being restored. Clearly the lateral coordination between the legs can be stronger than the transverse, as has been found for Geophilomorpha and some other arthropods (Parts 1, p. 547; 3, p. 131; 7, p. 391). The patterns of the gaits of Lepisma and Thermobia at their fastest are approximately (6.7 : 3.3) to (6.0 : 4.0) and always at a phase difference between successive legs of At the former pattern the gait is unstable with only 2 legs in contact with the ground at all moments, and the latter has six short periods of support by three legs and six by two legs, per pace (figure 8i). An advantage of using paired legs in opposite phase is an increase in the frequency and a reduction in the duration of the unstable moments with two supporting legs, compared with the jumping gaits of Petrobius, cf. figure 8i for Lepisma, with paired legs in opposite phase at gait (6.0 : 4.0) p.d. 0.33, with
37 EVOLUTION OF HISXAPOD LOCOMOTORY MECHANISMS 239 figure 8f for Yetrobius in the same pattern of gait but paired legs in the same phase. Maximum speeds of running in Lepismatidae have not been recorded. The animals are extremely fleet, and they run fastest when first disturbed, in sudden darts, stopping, resting or turning, before running off at variable speeds. They are probably quite as fleet: as the Machilidae, in spite of their much shorter legs. (figures 31, 34, 36). The stride lengths, < 3 mm for a 5 mm Thermobiu (pl. 5, figure c) and < 3 mm for an 8 mm Lepisma, not in its fastest gait (pl. 6, figure h), are long considering the length of the legs. Two stride lengths just exceed the body length (heavy white line on pl. 5, figure c) an achievement intermediate between those of the longer legged Petrobius recorded on pl. 5, figures a, b, and similarly marked. The pace duration in the Lepismatidae must be very brief at fastest running, probably of the order of 0.02 sec. or less. It is also probable that the leg movements of the Lepismatidae are essentially jumping or hopping movements, as in the Machilidae described above and see 5 7C(iii). Jumping movements and unstable positions are easy to maintain when the pace duration is very short. The Lepismatidae can run very rapidly, even on vertical surfaces. (vi) Conclusions concerning the Thysunuru The Machilidae and Lepismatidae thus have in common the exploitation of fast patterned jumping gaits of unstable nature, made practicable by very rapid stepping, in contrast to all other classes of hexapods. The differences between the Machilidae and Lepismatidae are not fundamental, although only the former utilize high-jumping escape reactions, while the latter accomplish fleetness and rapid turning by short legs which hardly project beyond the flanks, and cause no hindrance when moving in confined spaces. The Lepismatidae employ a leg movement (5 7C(iii)) which contrasts with that described in 5 4A for hexapods in general. The Thysanura break the man-made rules for hexapod running based upon the Pterygota (Hughes, 1952) in that legs 1 and 2 do not remain in contact with the ground until the footfalls of legs 2 and 3 respectively, and the paired legs do not necessarily move in opposite phase relationship. Presumably the ancestral Thysanura embarked upon the exploitation of fast, unstable jumping gaits, with patterns shifting from the stable (5.0 : 5.0) towards the unstable (6.7 : 3.3) gait, by developing short pace durations giving sufficient speed and momentum to carry the animal over the unstable moments. The opposite phase relationship between paired legs is presumably the primitive one, and is still retained by the Lepismatidae at most times and by the Machilidae when walking on ceilings of rock, etc. Throughout the arthropods described in previous Parts there are examples of animals in which the coordination between legs of a pair is not constant (Onychophora, Geophilomorpha, the scolopendromorph Cruterostigmus) and the Thysanura can be added to this list. Irregularities in the several sequences of unilateral longitudinal coordination are rare, the manner of stepping, described in 5 4A, requiring this. It is the longitudinal coordination which is the most stable and important, the transverse coordination can be more variable.
38 240 S. M. MANTON 6D. PROTURA The Protura are small in size, mostly < 1 mm, a few reaching 2 mm in length. There is little range in general form of the body. For the present work living animals have been obtained from mixed woodland litter by putting it in funnels so that the animals walk down as the litter dries. The gaits of Acerentomon nemorale (Womersley) are recorded below. P (i) Habits and habitats of the Protura The habitats of the Protura have been summarized by Kiihnelt (1961): the moisture must be moderate. The animals avoid dryness and also too wet conditions, and some organic content in the soil is necessary. They do not penetrate deeper than 10mm into the soil. Surface living species, with relatively longer legs, e.g. Acerentomon doderoi, Eosentomon spinosum, E. armatum, can be distinguished from the deeper forms with decidedly short legs, e.g. Protentomon minimus and P. clevedonense. Sturm (1959) observed Acerentomon piercing the hyphae of fungal mycorhiza growing on the roots of oak and hornbeam, and the restriction of this animal to slightly acid soils tallies with the distribution of such ectotrophic mycorhizae which grow only under slightly acid conditions. Eosentomon also feeds on mycorhizal fungi, but is not restricted to them, and has been found by Sturm to feed on various free hyphae. Thus no great agility, or speed of walking, is needed in pursuit of food and smallness in size together with living considerably under cover provides adequate protection from predators. (ii) The walking abilities of the Protura The Protura, as exemplified by Acerentomon nemorale, are slow walkers. An animal 1.8 mm long achieved 0.5 mm/sec. or less when walking on a comfortable damp flat surface. Pace durations are long, of the order of 0.4 sec. or more. Only legs 2 and 3 are used in regular stepping. Legs 1 do not move in any fixed phase relationship with the rest, or with each other. Legs 1 can step, sometimes together, sometimes alternately, by a movement of forward extension followed by flexure without lateral projection (pl. 1, figures i, j, k). During walking on a flat surface legs 1 are often held stationary and forwardly extended on either side of the head. Whether they are primarily sensory, or whether the tips when applied to the ground exert a propulsive drag on the body, pulling it forwards into narrow places without the hindrance of projecting elbows, is uncertain. When walking on an exposed surface legs 2 and 3 step rhythmically, the fastest recorded gait pattern being (2.0 : 8.0) at a p.d. of 0.25 (figure 2c). This gait gives footfalls at even intervals of time and three legs simultaneously on the ground, but with four brief moments of support by four legs (figure 2j, k). This gait is as stable as can be provided by four legs. Any faster pattern, i.e. reduction in the relative duration of the backstroke, would result in unstable moments with only two legs in contact with the ground. Such a performance, combined with the slow stepping, would be quite unsuitable, cf. Lepisma which
39 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 241 can manage unstable moments during the pace because of its very rapid stepping (see above). The gait pattern (2.0 : 8.0), when employed by three pairs of legs and using a phase difference between successive legs of 0.33, provides footfalls at even time intervals and even loading on the legs (figure 2a, d, g). But the Protura, using only two pairs of legs, can gain the same advantages only by decreasing the phase difference to 0.25 (figure 2c, j, k). Elimination of the use of legs 1 in figure 2d would result in uneven footfalls and be as undesirable as a hexapodous use of the gait in figure 2c, f. The contrast between figures 2f and j show the disadvantage to a hexapod and the advantage to a quadruped in the use of the gait shown in 2c. (iii) Conclusions concerning the locomotory achievements of the Protura The Protura contrast with most hexapods in that legs 3 are not used for walking. A walking quadruped, whether it be a horse or a hexapod, uses slow patterns of gait, so maintaining even footfall intervals and even loading on the legs. These gaits show two successive footfalls on one side of the body, the alternation of the two sides being absent. This loss of alternate footfalls on the two sides has been shown (9 4B(iii) and figure 4) to be the inevitable result of increasing the relative duration of the backstroke, which brings corresponding changes in the phase difference between successive legs and hence changes in the footfalls (figures 4a-j and 2j). Such footfall sequence is no special innovation by the Protura. When allowance is made for size, the Protura are by far the slowest walkers recorded here among the apterygote classes. The marked flexibility of the proturan trunk is presumably useful in soil penetration. 6E. CONCLUSIONS CONCERNING THE LOCOMOTORY ACHIEVEMENTS OF THE HEXAPOD CLASSES There is, of necessity, a measure of similarity between the gaits of all animals walking on only three pairs of legs, whether they may be closely related or not (see certain crustaceans and arachnids in Part 2 and the classes of the Hexapoda above). The gaits employed are largely of the slower types owing to stability requirements. There are striking differences between the locomotory achievements of the several hexapod classes, which are associated with predominant habits. Each class has its own manner of leg movement, often reflected in the tracks, and its own choice of gaits. Many of these features are dependent upon detailed morphology of body and legs and the very different types of coxa-body union (see ). The need for stability is overriding for the slow movers, but momentary instability, when the body is supported by only two of the six legs, is a normal feature of fast moving Thysanura and sometimes Campodea. The strong slow movements of the legs of Jupyx and the extremely rapid and wide movements of the legs of Lepismu are related to habits and are made possible by* remarkable morphology (5 7C). This includes the unique telescopic antennae of the japygids (5 6A(ii)) which assist the crevice crawling of these animals.
.")
40 242 S. M. MANTON Of particular interest are the unstable jumping thysanuran gaits, the variable phase relationship between legs of a pair, and the abdominal jumping escape reactions of the Machilidae ( 5 9C). Collembolan limb movements can only be appreciated after an analysis of the hydrostatic jumping mechanism by the spring organ, which has influenced the whole anatomy and movements of body and legs (5 9A, B). The component features of hexapod gaits are smoothly integrated so that the many gaits which are practised by these animals form even series controlled by known factors. Changes in the relative duration of the forward and backstroke, affecting the speed, are automatically accompanied by alterations in the phase difference between successive legs and in the order of footfalls. Even the loss of alternate footfalls on the two sides of the body is an obligatory result in the gaits at the slow end of the series. And, unless speed is a particular objective, even loading on the legs is as desirable for a hexapod as for a myriapod, but this asset is usually not possible when the fastest gaits are employed. 7. THORACIC PLEURON, LIMB MORPHOLOGY AND FUNCTION IN THE APTERYGOTE CLASSES The account which follows of apterygote morphology contains much new data, and throughout the approach is functional. Very little information concerning function has previously been recorded. Before embarking upon the detail it may be useful to note some of the many ideas and theories abounding in entomological literature which find no support in the present work. No evidence has been found which upholds the view that the Apterygota are a unified group of primitive insects. The fallacy of regarding the entognathous classes as closely united phylogenetically has already been given (Manton, 1964 and see 10G). The idea that the Apterygota exhibit progressive degrees of desclerotization (Matsuda, 1963) appears to have little foundation and no functional reasons for an initial high degree of sclerotization followed by an evolutionary reversal have been found. The idea appears to be one which takes no account of function or of functional continuity during evolution. The view that the leg-base, the coxa-body junction and associated pleural structures, can be described in common terms for the apterygote classes is untenable because no recognition has been given of the fundamental and mutually exclusive differences, both functional and morphological. No hypothetical generalised diagrams of ancesn-a1 states of subcoxal structures (e.g. Matsuda, 1960, 1963) can mean anything unless some functional explanation of their use can be provided, and this has never been done. The dorsal (tergo-pleural) leg-support in the Thysanura, the ventral (coxo-sternal) articulation in the Diplura and in the Collembola the entire lack of any coxa-body articulation, but with an elaborate internal suspensory system taking the place of an articulation (a feature related to the hydrostatics of the spring mechanism) are here shown to be fundamentally different from each other and also from the proturan (sterno-pleural) coxal support. The pleural armature of the apterygote classes, and the functions which the parts subserve, differ markedly from class to class. The leg of Lepisma ranks as
41 EVOLUTION OF HISXAPOD LOCOMOTORY MECHANISMS 243 one of the most specialized among hexapods in its structure and mode of action (57C(iii)), and the pleural arrangements here, so different for those of the Machilidae, cannot be regarded as basic for all other hexapods. The functions of pleural sclerotizations, of the subdivisions of collembolan coxae and of claimed subcoxae have been carefully considered, and different conclusions have been reached concerning their interpretation and existence. The locomotory and other achievements of the hexapod classes which have been described are dependant upon body structure which differs greatly from class to class. The correlations between the movements and the mechanisms are deep-seated. The whole trunk and limb morphology may be geared to the execution of one particular habit, such as jumping by the Collembola, all other habits being subservient. In the Collembola amazing structural modifications associated with jumping are all-embracing, the morphological details combining to provide a hydrostatic jumping mechanism with which the evolution of this class basically is associated. The similarities between the hexapod classes, although easily apprehended, are more superficial than are their fundamental differences. These classes must have followed their own independent paths of evolution, perfecting their morphology in association with their particular habit proficiencies. The legs of the Diplura and Protura project from the body ventro-laterally, as in Chilopoda, Symphyla and Pauropoda (figures 9, 12, Part 9, figures 4-8) in contrast to the ventrally directed coxae of the Collembola, Thysanura Machilidae and the Pterygota (figures 22-25, 3 1, Part 9, figure 10a). Distal to the coxa and trochanter the collembolan leg possesses two podomeres and the Diplura, Protura and Thysanura three podomeres, in contrast to the basic three or four in Myriapoda. In Part 9 it was suggested that the podomeres of the several classes of myriapods are probably not directly homologous one with another, and that a primitive, probably unsegmented, leg became sclerotized and subdivided differently according to functional needs. This conclusion appears to hold also for the apterygote hexapods ( 5 10E). In Part 9, p. 125, figure 10 the coxa-sternite articulation of a chilopod limb (ringed) was contrasted with the opposite arrangement in a pterygote where the coxa is principally supported by a coxa-pleurite articulation (also ringed). The two conditions are mutually exclusive, and both show wide coxal movement to take place on the opposite side to the articulation, dorsal in the chilopod and ventral in the pterygote, where much flexible membrane is present. The coxa-body junction of the chilopod in essentials is characteristic of other classes of Myriapoda. The apterygote classes each have their own type of coxa-body junction which differs from both the myriapod and the pterygote types- The Myriapoda show a series of distal hinge joints on the leg which lack extensor muscles (marked by unringed black spots in Part 9, figures 9, 10). Except for the intra-tarsal joints, the Pterygota usually depend on dicondulic pivot joints worked by antagonistic pairs of muscles (marked by ringed black spots). Their plantigrade, multisegmented tarsus shows considerable flexibility, giving a good grip on a variety of surfaces and at inclined angles. But the apterygote classes all possess a long strong unguligrade tarsus equipped with a proximal strong hinge joint, which, as in the Myriapoda, lacks extensor muscles. It has been shown in Part 6, text-fig. 8 and Part 8, p. 258 (figures 2, 3)
42 244 S. M. MANTON how a rocking of the leg base in the parasagittal plane about the ventral coxa-sternite articulation, brings the dorsal face of the leg to a more forward position during the remotor backstroke. This movement facilitates extension of the distal hinge joints, lacking extensor muscles, during the propulsive backstroke, and is effected by extrinsic and other muscles in Chilopoda, Symphyla and Pauropoda, but in the Diplopoda the advantage is gained by the set of the legs on the sternite. This same asset is obtained by other means in the apterygotes. 7A. THE PLEURON, LEG, LEGBASE AND ASSOCIATED MOVEMENTS OF THE DIPLURA The Diplura, in contrast to the Collembola and Thysanura, possess an extensive array of pleural sclerotizations separated by flexible membrane. This morphology is associated with two functions, the marked thoracic flexibility shown by the Diplura in association with crevice penetration (8 6A(i)), combined with strong support for the coxa-body articulation. The shape and insertion of the coxa on the body, so that the limb base projects laterally, as in Chilopoda, (figures 9, 11, 12, 14; Part 8, figures 46,47, 53, 54, 58, 63, 72, 73) is probably associated in both classes with crevice penetration for which the dorso-ventral span of the body and appendages is at a minimum. The pleuron of the Diplura superficially bears more resemblance to that of the Chilopoda than to any other class, but the resemblances are superficial, cf. figure 9 with Part 8, figures 41,48, 58, 65. The basic plan of the sclerotizations in the campodeids and japygids is fundamentally the same, although there are differences in detail. The japygids are much more strongly constructed; this tallies with their strong movements in the soil in contrast with the agility of the more lightly constructed campodeids, and the much larger size of some of the japygid species. The dipluran coxa shows the promotor-remotor swing on the body, as is usual in myriapods and hexapods. But there is no rocking of the coxa on the body as takes place in myriapods' (Part 8, figures 2, 3). A corresponding movement occurs in the leg itself (see below 0 7A(iii)). Levator-depressor movements of the coxa on the body are usually absent in myriapods, although contrived to a minor extent in the Symphyla (Part 9, figure 6). The levator movement of the myriapodan leg is dependent upon the coxa-trochanter joint, with all its specializations in hard parts, wide expanse of flexible arthrodial membrane and great difference in diameter of coxa and trochanter (Part 8, figure 58, p. 260). The Diplura contrast in possessing strong coxal levator muscles in both Campodea and Japyx (lev. co. 1, and in Campodea, where the coxa is more freely articulated, coxal depressor muscles (dep.co.) are also present (figures 12E, F, Zev.co., dep.co. ). More extensive levator and depressor movements occur at the dipluran coxa-trochanter joint, where there is considerable difference in 'diameter of the two podomeres, but not so great as in the Myriapoda (cf. figures 12E, F with Part 8, figures 58, 63, etc.). The dipluran leg is probably stronger than a myriapodan leg as a result of these differences in morphology and movements, as is suitable for an animal with fewer legs.
, but the sternal component of this articulation is differently")
43 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 245 (i) Coxa-sternite articulation, its movements and the extrinsic leg muscles of the Diplura The coxa articulates strongly with the sternite, as in the Chilopoda, Symphyla, Pauropoda and Diplopoda (Part 8, figure 58, Part 9, figures 6, 8e, lob, marked by black rings with open centres), but the sternal component of this articulation is differently contrived. It consists in japygids of the very strong anterior end of the arm of the Y-shaped cuticular ridge (infolding) which traverses the sternite. Externally this apodeme shows only as a slight groove, but internally it projects as a large flange, marked lateral arm of Y-ridge on figures 9 and 10 bottom left, and in section on figure 1 le, F, G. There is no coxal apodeme as in the Chilopoda, where this structure strengthens the complex coxal movements by providing suitable sites for muscle insertions (Part 8, p. 258). In the campodeids, where the legs are longer and the movements swift, the coxa-sternite articulation is as shown in figure 12E, F. A stout incurved cuticular flange at the ventral mesial margin of the coxa tucks under the edge of the sternite, but is linked with it by flexible membrane without an opposing sclerotization. But the sternite, so narrow at this point because the coxae sink into it towards the middle line, is very strongly supported internally by struts of cuticle which pass from the sternite to the lateral endoskeletal bar (figures 12E, EE, FF). These cuticular structures correspond with the arms of the Y-skeleton of japygids, although the details are different. The coxa-body joint permits promotor-remotor movements about a roughly vertical axis passing through the ventral coxa-sternite articulation (figure 9). Levator and depressor movements also take place on a small scale. The basic plan of the limb musculature of Citmpodea and Japyx is the same but the details are suited to their different needs, the legs of Campodea being long and capable of fast running, while those of japygids are short, stout and strong, and less suitable for surface locomotion. Extrinsic leg muscles extend from the proximal coxal rim, the marginal apodeme of japygids, and from the trochanter to the tergites, sternites and endoskeleton. Tlrere are no extrinsic muscles causing a rocking of the coxa, as present in myriapods. Protractor and retractor muscles. Coxal protractor muscles to the tergites in Cizmpodea form three or four groups. Muscle pr. co. 1. arises from the junction of the coxa and trochantin (figures 12E and 13b) and passes directly upwards to the tergite above. Muscle pr.co.2. arises more dorsally from the anterior proximal coxal rim and passes upwards, in two main sectors, to insert on the lateral and median parts of the tergite above. Muscle pr.co.3. leaves the proximal anterior coxal rim behind muscle pr. co. I., and passes backwards as a stout short muscle to the median apodemal endoskeletal plate (described in 5 8A(ii)) on the prothoracic segment (figure 12E). On the meso- and metathorax the corresponding muscle passes dorsally to the tergite (figure 12EE). Muscle pr.co.4., not present on the small prothoracic leg, arises from the anterior coxal margin and inserts on the lateral endoskeletal bar which is also apodemal (figure 12EE). In Japyx there are minor differences. A tergal protractor pr.co.2. in two sectors fanning fore and aft (figure 10, prothorax) corresponds with the similarly named muscle of Campodea, muscle pr. co. 1. being unrepresented.
44 246 S. M. MANTON Muscle pr. co.4. is short and stout, arising from the inturned coxal apodeme and inserting on a tendon alongside that of muscle dvc. at the coxa-sternite articulation, a point supported by the stout arms of the Y-skeleton (figure 11E). Coxal retractor muscles in Campodea comprise long, well-fanned-out tergal muscles; short, stout muscles inserting on the endoskeletal plate and its median union with the sternite; and a long muscle to the sternite. Muscles ret.co.l., ret.co. 2., ret.co. 3. leave the postero-dorsal coxal rim and insert on the tergite above, on the succeeding tergite, and on to the cranium from the prothoracic leg, as shown on figures 12 and 13. Just below these muscles arise ret.co.4. and ret.co. 5. passing to the median endoskeletal plate and sternite respectively, the latter crossing to the opposite side. Muscles ret. co. 6. comprise short stout sectors inserting on the posterior part of the median endoskeletal plate and its sternal origin from the cuticle (figure 12F, G). In Japyx muscle ret.co.1. inserts on the cranium, as in Campodea, but is not represented on the meso- and metathorax. Muscle ret.co.2., in one sector on the prothorax and two on the meso- and metathorax, corresponds with the similarly named muscles of Campodea, and ret. co. 3. is unrepresented. The endoskeletal differences between Campodea and Japyx account for the great superficial differences between the remaining retractor muscles. The enormous muscle complex, ret.co. 6. of Japyx, arising from an apodemal intucking in the arthrodial membrane behind the coxa, (see prothorax on figure 10) and inserting on the spine apodeme, corresponds with the similarly named muscles of Campodea, but leaves space for no others. The large sectional area of muscle ret. co. 6. (figure 11G) implies great strength and the great length, made possible by the spine apodeme, provides a stronger retractor system than possessed by Campodea or by any myriapod, and accounts for the strong retractor movements performed by the legs. Levator and depressor muscles. These muscles, absent in myriapods, are fairly well developed, but are not as bulky as the corresponding adductor and abductor muscles of Collembola. Muscle 1ev.co. arises from the dorsal rim of the coxa in both Japyx and Campodea. In Japyx this muscle inserts on the sternite between the arms of the Y-skeleton, crossing its fellow to reach the other side (figures 10, 11E, F). In Cumpodeu the muscle loops below the sternal retractor ret.co.5. of the coxa and inserts on the antero-lateral edge of a median endoskeletal plate (figure 12F). No myriapodan muscles of any kind have been found to loop round one another in this way. Contraction of the coxal retractor muscle ret.co.5. during the backstroke must stretch a relaxed Zev.co. muscle, and tend to depress the coxa, as is appropriate during this phase of the step. Contraction of Zev.co. can only take place effectively during the forward swing of the leg, or at other times when muscle ret.co.5. is not active. Slight levation of the coxa during the forward swing is an appropriate movement, and reduces the necessary levation at the coxa-trochanter joint. The difference in the insertions of muscles 1ev.co. in Gzmpodea and in Japyx is associated with the availability of convenient skeletal sites for attachment. The median endoskeletal plate of Campodea corresponds with the spinous apodeme of Japyx ( 8A), but is anterior enough in position to carry leu.co., the much smaller ventral expanse of sternite in Gzmpodea being amply used by other muscles and skeletal supports (cf. figures 10, 12E, F, FF).
45 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS An antagonistic depressor muscle to the coxa, dep.co., is present in Campodea, as shown in figure 12E. This muscle can only cause small displacements of the ventral coxal rim, permitted by the amount of arthrodial membrane at the coxa-sternite joint (figure 12E). In Japyx the amount of flexible membrane at the articulation is much less (figure 11E), no such movements are possible ventrally and there is no depressor muscle. Depressor muscle from the trochanter. This muscle, dep. tr. 2. fortifies the action of the intrinsic depressors to the trochanter (figure 12F). In Campodea the muscle leaves the proximal ventral margin of the trochanter and inserts on the median endoskeletal plate. The shortness of the coxa permits the presence of some extrinsic muscles to the trochanter, but not as many as in the Chilopoda where longer protractor, retractor, and levator muscles fan widely on to the large sternite. 247 (id Pleurites of the Diplura There are two major sclerotizations around and above the coxa in the dipluran pleuron, besides the regions corresponding with the myriapodan proand metacoxa. These are pleurite 1 or the trochantin of entomologists, and pleurite 2 the distal anapleurite of Barlet & Carpentier (1962). Both bear a row of large spines, those on the trochantin being drawn on figures 9a, 12E. These pleurites are separated by flexible pleuron, termed the catapleural zone or arc by Barlet and Carpentier, but since this region is not a sclerite, it is here considered to be misnamed (see 5 3). The coxal base is closely surrounded by the pleurites (figures 9, 12E, F), contrasting with most Chilopoda (Part 8, figures 48, 58, 74), the close investment in Geophilomorpha being correlated with the mode of burrowing. Dipluran pleurites support the protractorretractor swing about the axis marked on figure 9a, and inhibit movement in other planes. Pleun te Z, or the trochantin, closely surrounds the coxa in front, above and behind. There is no wide expanse of flexible membrane as there is between the coxa and the katapleure in scolopendromorph and anamorph Chilopoda which facilitates the rock (Part 8, figures 48, 58, 65, 74). There are functional reasons for the closer investment of the coxa by pleurites in the Geophilomorpha, and for the close association of coxa and procoxa in Craterostigmus (Part 8, pp. 262, 279, 317, figures 41, 73, 74) but these features do not hinder the rock. The trochantin of Japyx is a little more complex in form than in Campodea and makes no articulation with the coxa. An apparent point of close union antero-dorsally is the origin of the apodeme of the coxal protractor muscles overlapped by the trochantin (figures 9a, 11E, and in 12E, where the overlap by the trochantin is drawn as if transparent). The trochantin stabilizes the coxal movement about the single axis. Pleurite 2, situated as shown in figure 9a fades out postero-ventrally on the pleuron and is closely associated with the immobile procoxa anteriorly. Pleurite 2 is thus firmly fixed in position, so prohibiting any rocking movement of this pleurite. There are infoldings of membrane at the margins of pleurite 2, and the amount of exposed flexible pleuron between the trochantin and pleurite 2 depends upon the momentary position of the parts (cf. the pro- and mesothorax shown in figure 9a). The presence of some flexible pleuron is
46 248 S. M. MANTON necessary for he coxd promotor-remotor swing and for the n ~~ked ability of thoracic bending own by the Diplura (8 6A(i)). Pleurite 2 does not correspond either functionally or in position with the chilopodan katopleure but might be roughly likened to the chilopodan ano. The pro- and metacoxal sclerotizations, marke #feure On figure 9% correspond with the similarly named separate pleurites Of the Myriapoda (Part 8, figures 41,45a, 48, 58); the former is named merosternite in the Diplura by Barlet & Carpentier (1962). These sclerotizations in the Diplura are continuous with the sternite, a condition associated with the support they give to the coxa, helping to restrict its movement to the promotor-remotor swing on the body. As noted in previous Parts, there are many examples where skeleton, in restricting the movement at joints, enables a maximum of muscle to be employed in causing the most needful movements. The absence of any coxal rocking movement gives no need for the pro- and metacoxa to be moveable on the sternite, as they are in Chilopoda. The large and bulbous paired presternites and median intercalary sternite figure 9a) spread laterally and occupy the pleuron as far as the tergal sclerotizations. They provide strength combined with great thoracic flexibility in the transverse plane. Thoracic flexibility is prevented by a variety of means in Collembola and Thysanura. These sclerites do not move as do the intercalary sternites of Chilopoda (Part 8, p. 265). But by deepening the dividing furrows at their margins, body flexures result. The procoxal sclerite (figure 9a) is capable of overlapping the presternite to reach the intercalary sternite. The margins of pre- and intercalary sternites are a little less sclerotized than their middles, but there is no intervening arthrodial membrane, or zones of cuticle bearing cones of sclerotization, as in Geophilomorpha (Part 8, figures 8, 22, 23, p. 262). The firm furrow between the pre- and intercalary sternite prevents buckling and carries the well developed superficial dorso-ventral muscle (figure 10). In Campodea these sclerites are less sclerotized, but correspond with those of Japyx (Barlet & Carpentier, 1962, figures 1-3). The superficial dorso-ventral and superficial pleural muscles of the Diplura mediate the marked trunk flexibility, as they do in epimorphic Chilopoda and Symphyla, but these muscles do not cause any rocking of the leg base. This category of muscles is absent in Pauropoda where rigidity and not flexibility is needed (Part 9, p. 119). Campodea possesses superficial dorso-ventral muscles 1 and 2 (figures 12G, 13b) and a pleural muscle (marked so on figure 12E) which may assist lev. co. Japyx possesses three muscles marked: superficial pleural muscle, superficial dorso-ventral a and b on figure 10. Thus the pleural sclerotizations of the Diplura serve the needs of (i) strength and trunk flexibility and (ii) stability of the coxa for its promotor-remotor swing about its ventral coxal articulation. In particular the four sclerites, proand metacoxa, trochantin and pleurite 2 stabilize the coxa in contrast to the promotion of its mobility by 4 roughly corresponding sclerotizations in Chilopoda, the single pleurite of Symphyla and no pleurites in Pauropoda. (iii) Dipluran legs, their intrinsic rocking mechanism and intrinsic muscles As noted above, the coxa-body joint of the Diplura permits a promotor-remotor swing about a vertical axis (figure 9a) without the additional
47 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 249 rocking movement employed by many myriapods. Levator and depressor movements of the whole leg, as usual, are mainly served by the pivot joints at coxa-trochan ter, trochanter-femur and femur-tibia positions. The dipluran leg is unique in possessing a highly specialized femur-tibia joint and muscles which twist the femur at its proximal joint, so causing the distal parts of the leg to rock. The ventral face of the distal part of the leg is brought to a more posterior position, as is done by other means in the Chilopoda (Part 8, p. 258, figures 2, 3) where the dorsal face of the leg is rocked forwards. The result is the same in either case, facilitation of the extension of the tibia-tarsus hinge joint. The rocking movement is shown by pl. 2, figures a, c where legs 3 are in opposite positions. The left leg, near the end of its forward stroke, diplays the wide anterior face of the femur, the ventral face of the leg being rocked forwards, while the right leg, near the end of the backstroke, shows the narrower dorsal face of the femur, the ventral face having been rocked backwards. The rocking movement in japygids is less than in campodeids and the rocking musculature is less developed, as is appropriate where strength of legs, and not speed of running, appears to be a main requirement of japygids. The femora2 rocking mechanism. The trochanter joint is a modified pivot joint in which the anterior components imbricate strongly and there are no posterior articulations, so leaving the femur free to rock or twist about the anterior articulation within the loosely overlapping trochanter margin. Femoral rotator muscles r0t.fe.a and B pass as shown (figure 12F) from the ventral face of the femur and its proximal posterior rim, obliquely upwards to the proximal posterior rim of the trochanter. Contraction of these muscles pulls the ventral face of the femur backwards and upwards, so giving a slight twist to the distal part of the limb. The recovery movement of the femur is not mediated by any antagonistic muscles and the resting alignment, restored during the forward swing, is probably given by the tensions on many muscles and perhaps by some cuticular elasticity. A distal joint to the trochanter, such as just described, is not present in any myriapod and no muscles such as rot. fe. have been found elsewhere. The distal joint to the trochanter, if mobile at all, is a dicondulic pivot joint in Diplopoda, Pauropoda, Symphyla and Pterygota and it is a dorsal hinge joint in Geophilomorpha. But the homology implied by the term trochanter is probably not real (Part 9, pp ). The joint between the femur and tibia in the Diplura is also a derivative of a pivot joint and is constructed so as to transmit the rotation of the femur to the distal part of the leg. The anterior component is very strong indeed, and the femur is enlarged and prolonged into the process marked W in figure 14. There is no posterior component. The tibia is prolonged dorsally into the process marked Z from which arises the tendon bearing the levator muscles lev. ti. from the tibia (figures 12F, 14b, c). This process can only be seen externally when the tibia is very strongly depressed so that the shoulders marked X and Y are in the positions seen in figure 14b compared with (a). Thus, as the leg is extended during the latter part or during the whole of the backstroke in the case of leg 3, tension on muscle lev. ti. and close articulation between W and Y must transmit the rotation of the femur to the tibia, the proximal depressor muscles to the trochanter (and to the coxa in Campodea) keeping the limb tip on the ground.
48 250 S. M. MANTON The antagonistic depressor muscle to the tibia dep. ti. is long, as is muscle lev. ti., both inserting largely on the trochanter (figure 12F). As shown in Parts 6 and 8, long leg muscles can contribute considerable and rapid displacements of hard parts at joints, while short muscles, such as dep.tr., exert stronger but less extensive movements of skeleton. Leverage is also important. The hinge joint between tibia and tarsus is normal in its construction (figures 12F, 14d). Intn'nsic muscles. The principal levator and depressor movements of the leg take place at the coxa-trochanter joint, as in myriapods and pterygotes, but unlike the myriapods, small levator and depressor movements occur also at the coxa-body joint (see above). The coxa-trochanter pivot joint possesses strong anterior and posterior articulations and fairly ample dorsal and ventral expanses of arthrodial membrane, the latter being far less ample than in Chilopoda (cf. figure 12F with Part 8, figures 1, 63, 65, 73 and Part 9, figure 5). An anterior sclerotization in Japyx, comparable with the costa coxalis of Chilopoda, similarly strengthens the anterior component of the pivot joint. The dipluran leg is thus stronger than in Chilopoda, at the expense of the very easy levator-depressor movements of the latter. Antagonistic levator and depressor muscles from the trochanter, lev. tr. and dep. tr. pass proximally to the coxa as shown in figure 12F, the depressor being the larger and stronger muscle. The trochanter-femur joint is a modified pivot, as noted above in the previous section. Antagonistic muscles might be expected here if the pivot was normal but no such muscles are present. Instead there are strong rotator muscles which cross the joint (see above). Two sectors are present in Cizmpodea, r0t.fe.a and r0t.fe.b. In Japyx only r0t.fe.a is present and here telopod rocking is less well developed. The specialized femur-tibia joint (see previous section) and the antagonistic depressor and levator muscles dep. ti. and lev. ti. (figure 12F), both crossing two leg joints, have been noted. The tibia-tarsus hinge joint is very strong, a small flexor muscle, fl. ta. leaves the postero-ventral proximal margin of the tarsus and inserts proximally on the tibia. The tarsal claw consists of two strong hooks born on a common base which is hinged to the tarsus. The tendon from the tarsal claws bears very long and strong flexor muscles which insert on the tibia, femur and trochanter as shown on figure 12F. 7B. THE PLEURON, LEG, LEG-BASE AND ASSOCIATED MOVEMENTS IN COLLEMBOLA The jumping mechanism of Collembola has profoundly modified the entire morphology. A terminal springing organ on the abdomen projects the animal high into the air. Peculiarities of the cuticle, endoskeleton and tendons maintain a constant body volume by not permitting a billowing out of flexible, or infolded, cuticular regions under the momentary increases in internal hydrostatic pressure needed by the jumping mechanism (8 9A (v), (vi)). Tergo-pleural arches envelope most of the thorax, leaving very little pleuron exposed, in contrast to the Diplura. There is only one true pleurite and the rest of the pleural cuticle is considerably sclerotized and unstretchable. Alone among hexapods, the Collembola lack coxa-body articulations. At all
49 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 251 arthropodan hinge and pivot joints, and their derivatives, situated between coxa and body, or between the leg segments, there are imbricating sclerotizations separated by a short expanse of arthrodial membrane, which confer strength and localization of movement to the joints (Part 6, text-figs. 10, 14, 15, etc.). But none of these articulations can resist effectively a force which tends to slide the sclerites apart, such as the sudden increase in internal hydrostatic pressure which is associated with the collembolan jump. Special arrangements exist in Collembola at all these joints which prevent dislocation by such means, thereby stopping an increase in volume at the joints which would hinder the transmission of a momentary maximal hydrostatic pressure to the springing organ. The Collembola have played a large part in theories concerning subcoxae and their possible transformations, but there are no sound descriptions of these so called sclerites, or of the coxa-body junction. Denis (1949) writes On s accorde generalement i leur accorder deux anneaux su bcoxaux reccourcis du cote sternal. I1 n y a pas de Crete pleurale et la condition est tout i fait primitive. Les articulations sont tres ma1 dtfinies, ce qui est encore sans doute un caractere. primitif. Not one of these statements is substantiated here, and still less is the adjective primitive appropriate to the morphology. The conflicting views of previous workers on the leg structure of Collembola is also summarized by Denis (1949). The musculature of the distal part of the leg has been accurately described, but there has been no understanding of either the structure or of the functional significance of leg-base morphology. The only figure of leg musculature is that of Bomer (1921), showing an inaccurate representation of leg 1 with supposed subcoxae. An external lateral drawing of the trunk and appended legs of Tomocerus plumbeus is given by Carpentier (1949) but with his interpretation of the parts. The present findings of coliembolan leg morphology are described below and are shown in figures 18, 22-25, 28, 30. It is considered here that the entire coxa of leg 1 (marked ab on the figures) corresponds with the partially divided coxa of leg 3, (marked on the figures as regions a and b) and with the completely divided coxa on leg 2, where sclerotized regions a and b are separate. Functional and morphological reasons are given for this view (3 7B(iii)). It is also concluded that there is only one pleurite and that there are no subcoxae. (i) The coxa-sternite junction, its movements and the extrinsic leg-muscles in Collem bola The coxae do not project sideways from the body, as in Diplura, but hang down from it at slightly different angles (pl. 3, figures b-d). Stability is not maintained by articulating sclerotizations between coxa and body, but by an internal system of muscles and tendons which holds the leg-base in place. The thoracic tendinous endoskeletal system is shown in part in black on figures 18, 21a, 22-25, and a strong endoskeletal node marked G forms a point of suspension for the whole leg (figure 30). A dorsally directed process k is linked by muscles to the sub-ectodermal connective tissue complex elaborated at the site (figures 22B, 23E, 24H). The main endoskeletal complex lies in the ventral longitudinal muscular system, with branches to the deep dorso-ventral
50 252 S. M. MANTON and oblique muscles, and possesses many tonofibrilar ties to the sternal cuticle, marked on the figures (see also Appendix 11). On to the node G insert suspensory muscles from the coxa and from the coxal endoskeleton (see below), shown diagrammatically in figure 30. The leg-base and the telopod are thereby held in place, and cannot be blown slightly away from the body by any sudden increase in internal hydrostatic pressure at a jump, thereby ensuring no loss of trunk haemocoelic pressure at the moment when it is required to work the springing organ (see 3 9A). The proximal rim of the coxa is linked with the flexible pleuron and sternite, as shown in the figures. The degree of sclerotization of the coxal cuticle sharply contrasts with that of the surrounding cuticle (figure 16i, j) but the differences cannot be shown on the scale of the other figures. Leg 1 moves more freely on the body than do legs 2 and 3, as determined by manipulation of prepared cuticles and confirmed by photographs of the legs in action. A zone of least freedom on legs 2 and 3 lies at the antero-lateral coxal margin, and it is here that there is a link between the coxal face and a flexible tendinous strut r from the trunk endoskeleton (figures 23E, 24H). Leg 3 is also supported by an endoskeletal strut p (figures 24H, 28) and restricted in its lateral movements, apart from abduction and adducrion, by the pleurite. The needed flexures at the base of legs 2 and 3 are eased by the small intracoxal flexures made possible by the coxal subdivisions on these legs (see below). Any arthropodan joint lacking firm articulations shows a large number of muscles controlling its movements. The fairly free coxa-body joints of Chilopoda are controlled by up to 34 extrinsic muscles in contrast to the firmly articulating coxa-body joints of Diplopoda which are controlled by 2-4 muscles (Part 8, Table 2 p. 364; Part 6, text-figs. 4a, 7d, p. 529). Tomocerus possesses very many extrinsic muscles, numerous sectors of which are not separately described below. The extrinsic muscles serve two purposes, those causing movements of the coxa on the body and those suspending the coxa from the body in the absence of a coxa-body articulation. Suspensory muscles. As noted above, the endoskeletal nodes G are firmly held in the thorax and each lies above the internal cavity of a coxa. The leg is suspended from this node by muscle; which are evidently suspensory, rather than causing particular coxal movements, since their positions give little or no mechanical advantage. Muscles sus. 1. to sus.4. leave the coxal face and two muscles sus. co. pl. leave the unique endoskeletal coxal plate (see below, 5 7B(iv)) which spans the distal part of the coxa (figure 30). The suspensory muscles from the coxae insert on to the endoskeletal node G and the plate g behind it. The muscles are shown in figures 18a, 22-25, summarized in figure 30 and described in Appendix I. Protractor and retractor muscles. These comprise very many sectors and not all are separately drawn in the lateral view (figure 18a) or in the transverse sections (figures 22-25). Simple short protractors and retractors converge on to the endoskeletal node G, pr.co.e., ret. c0.e. and long protractors and retractors, in many sectors, pass upwards to the tergites and cranium. Muscles pro.co.a., pr.co. t., ret.co.a and ret.co. t. are considered to be serially homologous, as marked on the figures, and a muscles lie anterior to the t muscles and diverge from them to different extents. The sectors are elaborately arranged and not all in line with one another. Some additional promotor muscles arise: from the

51 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 253 proximal margin of coxal segment a on legs 2 and 3; pr.co., from the pleurite on segment 3 (marked pleural muscle on figure 18a) and from the pleuron above leg 1 as shown. On leg 1 the coxal promotor-remotor movements are greater than on leg 2, and for this reason and perhaps also because there is no firm exoskeleton above and in line with the resting coxa, the extrinsic protractor and retractor muscles diverge backwards to tergo-pleural arch 2, pr. co. t. and ret. co. t., and forwards to the head endoskeleton, pr. c0.a. and ret.co.a. The muscles pr. c0.a. and pr. co. t. from leg 2 still diverge a little but do not do so on leg 3, and similarly for muscles ref. co. a. and ret. co. t. on the two legs. But the protractor and retractor muscles as a whole from legs 2 and 3 form a converging group as they pass to the tergites above. This arrangement is in marked contrast to the prothorax and to other arthropods such as Chilopoda (Part 8, figures 44, 52 showing the spread to the sternite, Table 2, p. 364 lists the extrinsic muscles, their functions and the figures depicting them); the Pauropoda show a very simple dorsal spread (Part 9, figure 8) and similarly for the Diplura, figures 13a here. This unusual convergence of extrinsic muscles of legs 2 and 3 brings these muscles to zones above the endoskeletal nodes G about which each coxa moves, and provides a way of housing the many muscles required. That there are space difficulties in housing the meso- and metathoracic extrinsic muscles, as well as lateral longitudinals, is shown by the position of the former from leg 1 which pass internal to the lateral longitudinals, so gaining length by inserting high on the mesothorax, while extrinsics from legs 2 and 3 lie external to the lateral longitudinals and at progressively lower levels as the body width increases (figures 18a, 22-25). Abductor and adductor muscles. These are extremely well developed, and with the protractors and retractors, provide coxal movements in all directions on the body and also slight twisting on the long axis. Coxal adductor and abductor muscles are less well developed in Diplura and usually absent in Myriapoda. Abductor muscles leave the coxa laterally and insert 9n the tergo-pleural arch near to the extrinsic promotor and remotor muscles. One abductor, abd. leave coxa 1 (figure 22B). Three abductors leave coxa 2: the most anterior abd.1. from the face of the distal segment b, abd.2. from the proximal margin of coxal segment b, and abd.3. from the pleural fold between the coxa and the tergo-pleural arch (figures 18a, 23D, E). On leg 3 abductors abd.1. from the face and abd.2. from the margin of coxal segment b resemble those of leg 2. The third abductor abd.3. leaves the proximal margin of coxal segment a and is the most anterior of the three in position (figures 24G, H). The adductor muscles are more variously contrived on the three pairs of legs. Two wide adductors add. from coxa 1 pass from its antero-lateral face to the endoskeletal plate g on either side of the middle line (figure 22A). On leg 2 one similar muscle add. extends from the lateral proximal face of coxal segment b and inserts on the endoskeletal plate near the middle line, the insertion being supported by a fan of tonofibrils f (figures 23D and Dd showing the end of the adductor muscle just behind the general level of the reconstruction). On leg 3 the adductor movement is caused by the pleural muscle of that segment (figure 24H pleural mus. thorax 3 ) which arises from the sternal cuticle and from a tendon at the proximal coxal margin, and must swing the coxa inwards
52 254 S. M. MANTON about the flexible endoskeletal struts r and p. It is mechanically easier to cause adductor movements thus, when the outer part of the leg is supported by the pleurite and the endoskeletal struts. Depressor muscle from the trochanter. A single extrinsic muscle from the trochanter, dep.tr.l., arises from the deeper part of the common tendon from the proximal rim of the trochanter carrying muscles dep.tr. and dep.fe. The long muscle dep. tr.1. passes upwards to the endoskeletal plate g as shown on figures 22A, B, 23E, for legs 1 and 2. On leg 3 the muscle ends on the tendon shared with the pleural muscle of the metathorax which depresses (adducts) the coxa and therefore must assist the same movement of the trochanter (figure 24H). Extrinsic depressor muscles from the trochanter are uncommon. In the Geophilomorpha alone among Chilopoda a dep.tr.s. muscle passes to the sternite (Part 8, figure 46, Table 2, p. 364 and p. 281). The Diplura possess a dep. tr.2. muscle to the endoskeleton (figure 12F). (iil Pleurites and so-called subcoxae in Collembola Ventro-laterally, below the tergo-pleural arches of the thorax, the body surface is here described as pleuron. Potash preparations of the thoracic cuticle of jumping Collembola stained with chlorazol black, and serial sections stained with Mallory s triple stain, leaves no doubt that there is only one sclerite proper on the thoracic pleuron of these animals (figure 28). This pleurite lies as a lateral crescent round the base of leg 3, fading out postero-laterally and curling to a point round the antero-lateral part of the leg base. This unit has been termed the katapleure by Carpentier and others. But the pleuron above and anterior to it is no anapleural sclerite (Carpentier, 1949, figure 4 ap). It is simply the rest of the pleuron which extends upwards to the intersegmental groove between the meso- and metathorax, marked pleuron on the parasagittal section shown in figure 18a, just above and in front of the cut pleurite, which is also marked. The difference in the sclerotization of pleuron and pleurite cannot be shown on the scale of figure 18. The identification of the superficial structures is not at all easy, as shown by the scanning electron micrograph in pl. 4, figure b and the tracing of the same in figure 15d. The huge scales covering the cuticle and made obvious by gold palladium deposition, mask the limits of the sclerites. The pleurite pl is shown also in pl. 4, figure e at a higher magnification. Laterally above the pleurite the pleuron forms a line of flexible membrane between the pleurite and the tergo-pleural arch. Above the clearly defined sclerotized coxal cuticle of legs 1 and 2 lies an expanse of flexible pleuron, greater above leg 1 than above leg 2. On the mesothorax the pleuron forms a narrow flexible zone between the tergo-pleural arch and coxa, marked pleuron on figure 23F (a denoting the proximal coxal segment). It shows also as a small cuticular expanse on figure 18a above and in front of coxal segment a and passing towards the ventral sternal intucking giving origin to connective 10 (marked). The pleuron above leg 2 cannot be seen in the electron micrographs because of the overlap of scales (pl. 4, figure b). Above and in front of leg 1 is a much greater expanse of pleuron which passes insensibly into the flexible sternal and neck cuticle (figure 28).
53 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 255 Structurally this pleural cuticle is quite unlike the sclerotized cuticle of the coxa (figure 16i, j, 1 and see below). The pleuron cuticle is undifferentiated and falls into the bulges shown by the micrograph pl. 4, figure b and figure 15d. Three bulges are shown in section on figure 18a, marked pleuron above and to the left of leg 1, and the same bulges are cut transversely in figure 22A. The latter shows that at the marked points 1, 2 and 3 endoskeletal struts are anchored to the cuticle and thus hold it in, but permit the bulging in between. Struts 2 and 3 on figure 18a insert on the cuticle in a more lateral position than the plane of the reconstruction. The identification of the two bulges of pleuron nearest to the coxa of leg 1 as two subcoxae seems to rest on Borner s single inaccurate figure (1921, fig. 14), and so we now lose one of the foundation stones of the entomological edifice concerning subcoxae. The function of the endoskeletal struts is to anchor the prothoracic endoskeleton, and to prevent undue billowing outwards of flexible pleuron under increase of internal hydrostatic pressure. These bulges are labelled trochantin by Carpentier (1949), but as noted above, they are not sclerotized pleurites at all, still less are they subcoxal segments. The flexible membrane surrounding the coxae permits mobility of the leg upon the trunk, and that of leg 1 is the greatest. (iii) Coxal subdivisions and their functional and theoretical significarice in Collembola Serial sections stained with Mallory s triple stain and preparations of cuticle stained with chlorazol black, clarify the details of the coxal subdivisions. A consideration of their muscular connections and functions leaves no doubt that these are purely coxal structures and no support has been found for the view that they represent subcoxal units or pleurites, etc. The coxal cuticle on all legs is similar in structure and brown in colour owing to sclerotization. The flexible pleuron above the coxae is not brown. In sections stained with Mallory s triple stain the coxal cuticle appears as shown in figure 16i, in contrast to the pleural cuticle in 161. The coxal cuticle in the region normally occupied by very flexible arthrodial membrane between coxa and trochanter is thinner than elsewhere on the coxa (figure 22, cf. 16i), well sclerotized externally and with lesser sclerotization below. The coxa of leg 1 is undivided and marked ab on figures 15d, 18a and 22, figure 15d being a tracing of pl. 4, figure b and similarly marked. On leg 2 the coxal cuticle is divided by a precise ring of lesser sclerotization into a proximal segment a and a distal segment b (figures 15d, 23, 28, pl. 4, figure b). The proximal segment is short and shaped as shown. On leg 3 the division between segments a and b is incomplete. The limits of the flexible membrane between them is shown in figure 24, it forms a lateral crescent, the coxal cuticle being entire on the median face. Such slots are of frequent occurrence in arthropods, e.g. scorpion at the base of the trochanter (Manton, 1958a, figure 5B) and at the base of the manubrium in Tomocerzw, figure 17a, c, places where extra mobility is required. The musculatures of legs 2 and 3 are essentially similar, and there appears to be no reason to suppose that coxal segments a and b on leg 2 do not represent the partially divided segments a and b on leg 3. On both legs the proximal coxal
54 256 S. M. MANTON segment is supported by the endoskeletal strut r and the single pleurite above leg 3 is supported by the strut p. Carpentier (1949) regards coxal segment a, the proximal segment, as a metamorphosed pleurite, the trochantin, a conclusion not supported here. He homologizes the entire coxa of leg 1 (marked ab here) with the distal segment, complete and otherwise on legs 2 and 3, identifying the pleural bulge above the first coxa as a trochantin. The present work supports the view that the entire coxa of leg 1 (ub on the figure) corresponds with the partially divided parts u and b on leg 3, and that functional considerations account for the differences in coxal structure on the three pairs of legs. The undivided median side of the 3rd coxa corresponds exactly in form and musculature with the same on the 1st coxa (figure 18a for leg 1 and 24H for leg 3). The mechanical significance of the undivided, divided and partially divided coxae on legs 1-3 appears to be twofold. Complete or partial division will permit easier adductor-abductor movements in that the displacement can be shared between the coxa-body and intra-coxal joints. This movement is needed most by the second legs, and in them this joint is complete. The third legs need greater strength, gained by the solid median coxal face and the supporting pleurite. The coxae of the first legs are so freely inserted on the body that a subdivision of the coxa is needless. Secondly, the length and disposition of the extrinsic leg muscles is greatly affected by the form of the coxae. The bulk of the promotor and remotor muscles arise from the proximal coxal margin of leg 1 and from the proximal margins of the distal coxal segment b on legs 2 and 3 (figure 18a). If these muscles arose from the proximal margin of segment a on the second and third coxae, the muscles would be very much shorter, so that the same degree of muscular connaction would cause a much smaller displacement of the coxae. Thus length of muscle is clearly advantageous. But on leg 1 length is obtained in another manner. The extrinsic muscles do not converge, as they do above legs 2 and 3. They fan out to the second tergo-pleural arch and to the head endoskeleton (figure 18a) and equal the lengths of the extrinsic muscles of legs 2 and 3, so that a coxal subdivision on leg 1 is needless for increasing extrinsic muscle length. It is probable also that the convergence dorsally of the meso- and metathoracic extrinsic muscles may have been made practicable by the intra-coxal flexibility on xhesesegments, the convergence itself being a way of housing the many muscles required without interference with one another, but also permitting the thoracic segments to be shorter than in the Diplura (cf. figures 9a, 10, 13 with 18, 20 etc.). This and the fusion of the pro- and mesothoracic tergites aids the maintenance of trunk rigidity. Thus no support is found for the view that the proximal coxal segment of legs 2 and 3 are something other than coxal in nature; they do not appear to be modified trochantins appertaining to the pleuron proper, and no trochantins are present. The subdivision of coxae, partial or complete is explicable on simple direct functional needs. (iv) Collembolun legs, their suspensory system and intrinsic muscles The legs are usually strongly flexed at the coxa-trochanter joint, so that each nochanter and femur project roughly horizontally outwards from the vertical

55 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 257 coxa (figures 29, 30). A pivot joint between trochanter and femur permits small movements to be caused by antagonistic muscles, but the two leg segments are usually in line with one another. A trochanter-femur joint sometimes has been found to be absent on one or both of legs 2 and 3 (figure 29b, c) and Borner (1902) records a similar abnormality on leg 1 of one side of the body. The femur-tarsus joint is a strong hinge dorsally with much emargination on the opposite sides of both podomeres, allowing the tarsus to flex as shown in figure 29a. The distal part of the leg has one segment less than the legs of Pterygota, and fewer segments than found in any myriapod of other apterygote class. Since every leg-joint must be able to resist dislocation by sudden increase in hydrostatic pressure at a jump, it is not surprising that there are as few joints as possible, and that stability measures exist at all the joints (see below). The suspensory system in the legs of Collembola is unique among the arthropods so far investigated. Its presence is associated with the absence of a coxa-body articulation, and with the need to prevent the leg base, and the more distal leg joints, from being blown out and dislocated by sudden increases of internal hydrostatic pressure at a jump. The coxa-trochanter and trochanter-femur joints are simple pivot joints (see Part 6, text-fig. 10) with weak anterior and posterior points of close union between the stiffer parts of the podomere cuticle. Collembolan strength at the pivoting points is not obtained by strong imbricating cuticular features, such as characterize the diplopod and chilopod legs (Parts 6, 8), and such structures would be of little assistance in resisting internal pressures. Instead a series of internal guy lines has been evolved, summarized in the diagram, figure 30. In the distal part of each coxa lie a tendinous coxal plate, set antero-posteriorly across the lumen. Figure 18a shows the plate, marked, in surface view on leg 2. It is anchored to the cuticle by tonofibrils. When the coxa is cut in a plane at right angles to this, the plate is cut transversely, as shown in figure 18a leg 1 (marked), and leg 3 is almost a mirror image of leg 1. Each coxal plate gives origin to two muscles sus.co.pz. which lie anterior and posterior to one another. The anterior muscle inserts on the node G of the thoracic endoskeletal plates or on to a process therefrom, and the posterior muscle inserts more posteriorly on the endoskeletal plate from legs 1 and 2 but on to the node G from leg 3. This suspensory system aided by the muscles sus. 1. to sus.4. (see 8 7B(i)), strongly holds the coxae to the stabilized nodes G about which each moves, and prevents the coxae being blown away from the body on increase of hydrostatic pressure at a jump. The coxa-trochanter joint is also stabilized strongly in meeting the same need. A normally constructed coxa-trochanter pivot joint is very mobile, the close points of articulation are small and are situated on the middle of the anterior and posterior face, the anterior being the stronger (Part 6, figure 10, Part 8, figures 2, 5, Part 9, figure 5). These articulations are suited to control the plane of the levator-depressor movement of the trochanter, and they are not suitable for resisting sudden increases in internal hydrostatic pressure. The enormous costa coxalis apodeme of the Geophilomorpha serves, among other purposes, the stabilizing of the coxa-trochanter joint at moments of increase of hydrostatic pressure when burrowing, although this was not pointed out in Part 8.
56 258 S. M. MANTON In the Collembola a stout rope-like trochanter tendon spans the proximal trochanter lumen dorso-ventrally, situated near the anterior face (figure 18a, legs 1 and 3, but omitted on leg 2, figures 22B, 23E, 24G). Ventrally this tendon is connected by a fan of long tonofibrils to the cuticle and dorsally a narrow connective anchors the tendon to the proximal rim of the trochanter. The tendon carries substantial suspensory muscles to the trochanter, sus. tr. which fan widely on to the coxal faces, the several sectors not being exactly the same on all legs and are not all seen in any one view (figure 18a, leg 1, and figures 23E, 24G). The pull which these muscles exert must be largely suspensory in function, the mechanical advantage respecting the coxatrochanter pivot joint, which favours levator movement, is small. The levator to the femur, lev. fe., inserts on the distal side of the trochanter tendon. A fibrous strand leaves the coxal plate and passes distally down the lumen of the leg. From it branches pass to the anterior component of the coxa-trochanter pivot, to the trochanter tendon and to the anterior component of the trochanter-femur pivot. The fibres stain pale blue or purple and show no cross striations. From the posterior components of these two pivot joints, muscles ref. tr. and ret. fe. extend proximally as shown in figures 18a leg 1 and figures 29, 30 respectively. Muscle ret. ta. arises from the postero-dorsal margin of the tarsus close to the hinge articulation, it is a very large long muscle inserting on the proximal posterior part of the trochanter. This muscle can cause no appreciable tarsal movement and is clearly a stabilizer, preventing the femur-tarsus joint from being blown out and dislocated under increases of internal hydrostatic pressure. Thus these three successive telopod joints are most strongly suspended from the body. And in addition, there are a number of small muscles which prevent any blowing out of the necessarily large expanse of arthrodial membrane between coxa and trochanter, see mus.a on figures 23D, 24G and mus.b. on figures 23F. A summarising diagram of the suspensory system is given on figure 30. Intrinsic muscles. Intrinsic muscles, additional to those already mentioned in the previous section, are simple. The proximal margin of the trochanter gives origin to the normal antagonistic pair which work the pivot joint between coxa and trochanter. The levator muscle 1ev.tr. arises from the dorsal margin and spreads a little way along the root of the trochanter tendon (figure 18a, leg 1 unmarked, leg 2 marked, and figures 23D, 24G, H). The depressor muscle dep. tr. arises partly from the ventral rim of the trochanter (figure 24G, H) and a substantial connective tissue flange, projecting inwards from the margin, provides sites for the origin of large deep sectors of this muscle (figure 18a legs 1 and 3, 22B, 24G, H). The distal face of this flange bears the depressor from the femur dep. fe. The trochanter-femur joint is a simple, not very mobile, pivot joint, the anterior articulation being marked on figures 29a, 30. The joint is worked by the antagonistic pair of muscles lev. fe. and dep. fe. already mentioned. When the joint is abnormally absent (see above) sometimes the normal depressor, retractor and levator muscles arise from the undivided femur-trochanter cuticle in their normal positions (figure 29c). But in other cases these muscles, as well as the joint, are absent (figure 29b). No clearer example could be found within the same species of all stages between a functional joint and its degeneration. The femur-tarsus joint is a simple hinge joint possessing the usual strong
57 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS flexor-muscles. Short muscles,.fl. fa. pass from the ventro-lateral margin of the tarsus to the dorsal face of the femur, as shown on figure 29, and a long muscle fl, ta.i., median in origin, passes forwards to the trochanter. The muscle ret. ta. (figure 29) arises close to the dorsal hinge articulation and therefore has practically no leverage for causing flexures. The muscle cannot be the antagonist of fl.ta. and fl.ta.l., as might be inferred without consideration of the nature of the joint. Muscle ret.ta. is a stabilizing muscle, and, as already noted, the three muscles ret.tr., ret.fe. and ret.ta. form part of the system of structures which prevent dislocation of the leg by sudden rises in internal pressure. The tendon from the tarsal claws gives origin to the usual strong flexor muscles fl. urz. inserting on the femur and tarsus (figure 29) C. THE PLEURON, LEG, LEG-BASE AND ASSOCIATED MOVEMENTS IN THE THYSANURA The pleuron, leg and leg-base of the Machilidae and Lepismatidae are superficially so unlike as to need independent descriptions. Yet both are based upon common features, the details of which are correlated with the unique jumping gaits of the Machilidae and the equally unusual running in the Lepismatidae by a very rapid movement of the distal part of the leg. A key to the understanding of the evolution of the thysanuran pleuron and legs lies in the mechanism providing rapid and strong coxa-trochanter movements, such as exists in no other class of hexapods, and has enabled the proficiencies of both Machilidae and Lepismatidae to have arisen. In Petrobizis the coxae are long and pendant from the body (pl. 6, figure a, leg 2 and figures 3 1, 32), the telopod being strongly flexed at the coxa-trochanter joint. At rest the coxae of legs 1 and 2 are roughly vertical while those of leg 3 are usually backwardly directed (pl. 6, figures a, d; figure 3 lb). In the interests of brevity no qualification is given below respecting these differences between the three pairs of coxae. Two types of jumping are employed, the normal jumping gait in which each pair of thoracic legs takes off in sequence, and the large escape jumps, high into the air, which are effected by the abdomen. In Lepisma the stance is different. The legs are shorter and the horizontally flattened coxae are backwardly directed at all times, forming wide plates below their own, and part of the following, segment (figures 34, 35F, G, H). Each coxa of Lepisma works horizontally in a deep groove between the underlying flange from the sternite and the paratergal lobe, figure 35. The stepping movement is unique, it is largely restricted to the telopod where the levator-depressor movement at the coxa-trochanter joint becomes the propulsive (backward) and recovery (forward) swing of the foot. The Machilidae and Lepismatidae possess only one coxa-body articulation. It lies with a pleurite, which derives its support from the tergite. The Thysanura thus stand in marked contrast to the single sternal coxa-body articulation of the Diplura and the endoskeletal support of the coxa in the Collembola where the coxa lacks any proximal articulation. Barlet (1951, 1953, 1954, 1967) has provided a wealth of descriptive detail of muscles, exo- and endoskeleton of a machilid and a lepismatid. Much of this detail is confirmed by present findings, and reference is made below to the principal components of the musculature and associated skeleton, using the
58 260 S. M. MANTON muscle numbers of Barlet as far as is convenient. However the functional significance of the muscles is stressed below, because a proper understanding of the skeleto-muscular systems cannot otherwise be obtained. Tiegs (1955) described the pleural and extrinsic limb muscles of Ctenolepisma Zongicaudata, giving their functions as far as he could apprehend them. (i) Pleurites and their muscles in the Thysanura The pleuron of the Thysanura has been described in considerable detail by Barlet (1954, 1967), but, as for the other apterygote classes, it is considered here to be more serviceable to restrict the names of sclerites to actual sclerotizations, and not to give these names to areas, flexible or otherwise, mapping the whole pleuron (see 9 3). In Petrobius there is only one pleurite on the meso- and metathorax, the catapleurite of Barlet. It is horizontally placed. Anteriorly it gives origin to a large apodeme which lies over the coxal cavity and ends posteriorly below the edge of the endoskeletal plate (figures 32D, E, F). The single articulation with the coxa lies just anterior to the origin of the pleural apodeme, but the link of flexible membrane between the outer proximal rim of the coxa and the length of the pleurite (figure 31b) forms in effect a strong antero-posterior hinge which permits free adductor-abductor movement of the coxa on the pleurite. The anterior end of the pleurite is strongly supported. It is linked by very little flexible cuticle to sclerite Sa of Barlet (1950, 1967) on the underside of the paratergal lobe. This sclerite is amply supported by tonofibrils passing through both layers of ectoderm, so uniting sclerite Sa with the outer cuticle of the tergite. These fibres are contained within the ectoderm, they are unstriated and typically tonofibrillar in form and cannot represent a muscle, as claimed by Baslet, they are marked 117 on figure 32C. The fibrils slope outwards and downwards and must brace the pleurite against the pull of the protractor muscles 113, 114. The anterior end of the pleurite is also supported in the opposite direction by a large cone of tonofibrils projecting forwards and upwards to end on the basement membrane of the tergal ectoderm (figure 32B). Elsewhere the pleurite is unsupported, and its flexible link with the underside of the paratergal lobe becomes more ample in the posterior direction, as shown by figure 32D-G. Sclerite Sa also extends posteriorly a little way, but only anteriorly is it conspicuously supported by tonofibrils 117. The protractor-retractor movement of the leg on the body takes place between the pleurite and the body on the meso- and metathorax. The pleurite swings from the sclerite Sa, the posterior end of the pleurite being pulled into the body during the backstroke, the ample flexible cuticle around it permitting this movement (see below 5j 7C(iv)). Apart from the invasion of the anterior end of the pleurite on meso- and metathorax by coxal protractor muscles (see below), probably originally located entirely on the coxa, there are few pleural muscles. A single small muscle 116 (figure 32E) leaves the dorsal side of the pleural apodeme and passes outwards to the tergite. It is probably a stabilizing muscle, the free end of the pleural apodeme being well anchored ventrally, so preventing any participation by the pleurite of the adductor-abductor movements of the coxa. On the prothorax of Petrobius there are two horizontal pleurites, marked X
59 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 261 and Y on figure 32A, set at an angle to one another. The upper pleurite X, the superior anapleure of Barlet (1967), extends forwards anteriorly (figure 3 lb) and ends on the sclerotization of the head cuticle at the external opening of the posterior tentorial apodeme (Manton, 1964, figure 22). Posteriorly pleurite X ends below the posterior extremity of the paratergal lobe of tergite 1. Pleurite Y, lies parallel to pleurite X. Anteriorly it fades out just posterior to the extremity of pleurite X and posteriorly it ends on a vertical rod of highly sclerotized cuticle situated at the extremity of the paratergal lobe and united with an arm of the endoskeletal tendon system. The latter is anchored to the cuticle by tonofibrils immediately ventral to the sclerite. PleuriteY forms a complex articulation with the coxa (see below). The flexible membrane between pleurites X and Y is not extensive, it forms a hinge line between the two. Reference to figure 32A shows that a bending of pleurite Y on X will ease an abductor movement, so lessening the necessary flexure at the coxa-pleurite articulation. Since one or other pleurite is fixed fairly firmly both anteriorly and posteriorly, there can be little movement of these sclerites on the body, other than the folding just described. The coxa thus depends largely upon movements at its pleural articulation. In Lepisma there are three pleural sclerites on the meso- and metathorax, as shown by the reproduction of Barlet s 1951 drawing, figure 33 of the mesothorax only, with a diagram of Barlet s pleural areas. There is some simplification on the prothorax. The areas, or folds, in which the pleural cuticle falls are only sclerotized markedly in parts (figure 35a, b, d) and the pleurites have considerable powers of overlapping each other (see the ventral parts of the anapleure and trochantin in figure 35a). The trochantin lies close to the coxa and articulates with it at articulation b (figure 35d). This pleurite is expanded below the coxa, as shown by the same section. The katapleure forms a vertical plate and is separated from the trochantin by plenty of fexible cuticle (figure 35a, d). The katapleural sclerite does not extend postero-dorsally behind the coxal articulation. The fold of pleuron here is little sclerotized (figure 35b). From the dorsal extremitf of the katapleure a pleural apodeme extends inwards, as shown by figure 35d level H. The apodemal sclerotization forms the floor of the intucking. In Petrobius the prothoracic coxal apodeme shows its heaviest sc!erotization in a morphologically similar position (cf. figure 32A). The pleurites of Lepisma are all more mobile and possess more muscles than the single meso- and metathoracic pleurites and the two prothoracic pleurites of Petrobius. Pleurite movements in Lepisma are probably small, but very important in giving support to the rapid movements of the coxa and telopod. The internal end of the pleural apodeme of Lepisma is well anchored by muscles corresponding with the simple muscle 116 of Petrobius. Two substantial muscles from the ventral side of the apodeme (figure 35dH) pass laterally to the tergite at the origin of the paratergal lobe. Others pass from the internal end of the apodeme to the same situations, but also extend over the connective tissue sheet spanning the origin of the paratergal lobe, an abundance of large tonofibrils anchoring the latter to the paratergal cuticle, as shown. The internal end of the pleural apodeme is well supported by muscles 163, 164 of Barlet (1954) to the tergite above. In the opposite direction the internal end of
60 262 S. M, MANTON the apodeme carries the long extrinsic levator muscles from the trochanter, lev. tr.l., (see below). Barlet has described a series of muscles, , from the anterior margin of the katapleure to the tergite. The anapleure is a strongly sclerotized plate lying vertically in front of the katapleure and extending below it in a ventro-posterior direction figures 33, 3 5a, d, where it overlaps the trochantin. Part of the levators from the trochanter lev. tr.z.an. insert on the anapleure (figure 33c, d, e, see below). Barlet (1954) describes muscles 177, 178 leaving the anterior border of the anapleure to insert on the tergite, and muscles 179, 180 leave the pleuron more dorsally and also insert on the tergite. Tiegs (1955, Figure 1) describes and illustrates three tergo-pleural muscles of Ctenolepisma longicaudata, t.scx. 1., t.scx. 2., t.scx. 3. and one sterno-pleural muscle s.scx. The pleurites of Lepisma thus contrast with those of Petrobius in their equipment of muscles which must indicate greater mobility. Some of these muscles to the tergite (figure 3 5a) probably represent coxal protractor muscles (see above) which have shifted their origins from the coxal margin to the pleurites, as in Petrobius (figure 32C). As described below in $ 7C(ii), (iii), the protractor-retractor movements of the coxa of Lepisma are largely merged with the adductor-abductor ones, a great contrast to Petrobius where these two sets of movements on legs 2 and 3 are entirely distinct, as is usual in arthropods, and are provided for by the pleurite-body junction and by the coxa-pleurite junction respectively, but in a manner contrasting with all myriapods and other hex ap ods. (ii) The coxa-pleurite junction and its movements in the Thysanura In Petrobius rnaritirnus and Lepisma saccharina there is only one true coxal articulation with the trunk, in spite of statements to the contrary. Whether an articulation is present or not is shown precisely by sections stained with Mallory s triple stain or with Azan, in which the extent of sclerotization is plainly shown by the disposition of blue, red, orange and amber colouring. The position of this articulation is shown for Petrobius by black spots on figure 3 lb and for Lepisma by the ring marked articulation b on figure 3 3b. In Lepisrna a coxa-trochantin articulation lies between the dorsal rim of the coxa and the postero-dorsal margin of the arc-shaped trochantin. In section (figure 35d) a typical short arthrodial membrane separates the opposing sclerotizations. The solid direction line to the trochantinal sclerite shows that it forms only a portion of the pleural area of Barlet (195 1) bearing that name. The flexible part of this area is little sclerotized and is marked by a dotted direction line. The axis of the promotor-remotor swing of the coxa in Lepisrna passes obliquely through articulation b, as indicated by the heavy line added to the reproduction of Barlet s drawing, figure 3 3a. Ventrally this axis passes through the ringed point, named by Barlet articulation a. This is also cut in section in figure 35d. Here the sclerotized coxal cuticle is reflected at its ventral margin, as shown. The outer surface of the coxal cuticle does not articulate with the outer surface of the sclerotized trochantin. These parts are united by a wide fold of flexible cuticle. The reflected edge of coxal cuticle is separated from the trochantinal sclerotization by a double layer of ectoderm and internal body
61 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 263 space. Such an arrangement does not constitute an articulation at all, although the reflected margin must stiffen and strengthen the coxal margin in this region. The third coxa-body articulation of Barlet (1951) situated at the dotted circle just posterior to articulation b, does not exist. Level G in figure 35e lies just posterior to articulation b; beyond the coxal rim there is only the little sclerotized flexible cuticle marked trochantin and katopleure. These bulges of cuticle are continuous with the sclerites situated more anteriorly and bearing these names (figures 33b, 35a, d). No articulation is possible between the coxal margin and such little sclerotized cuticle. Flexible pleuron is seldom flat, preformed creases and lines of bending are present and maintained by fine cuticular structure, cuticular elasticity, etc. Folding may be conspicuous, but does not constitute an articulation. Owing to such surface features the planes of easiest movement are often delimited without further cuticular specialization. The pleural markings of scolopendromorph centipedes, for example, are too complex to be drawn with ease and an indication only is provided in Part 8, figure 1 etc. The coxal promotor-remotor swing in Lepisma is of limited extent, marked by arrows on coxa 2, figure 34. Owing to the position of the coxa, the movement is one of apparent adduction and abduction, almost in the horizontal plane, the coxa working between the expanded sternite below and the lateral extension of the tergite. Each thoracic sternite is wide anteriorly (figures 34b, 35a). The sternite slopes ventrally as a free flange, lying close to the coxae, as shown by figures 35a-c in which the body is drawn at the same horizontal level, so recording both coxa 2, and sternite 2 in descending positions. This remarkable backward projection of thoracic sternites, also shown in figures 33a, serves to support the very rapid promotor-remotor movements of the coxae. The three vertical pleurites just anterior to each coxa probably become less imbricated during the remotor movement, while still supporting the rim of the coxa; posteriorly the coxal rim deforms the pleuron, as in the remotor movements of chilopods, machilids, etc. In Petrobius the coxa-body articulation of legs 2 and 3 are similar and each is situated between the proximal anterior margin of the coxa and the single pleurite. On leg 1 the articulation lies between the dorso-lateral rim of the coxa and the middle of the lower of two pleurites (figure 31b). Figure 32C shows a section passing through this articulation on leg 2, where a very short link of arthrodial membrane joins the imbricating facets of thickened sclerotized coxal and pleurite cuticle. Elsewhere the coxa and pleurite are separated by a considerable expanse of flexible pleuron, shown in figure 32D-F, unlabelled, but situated between the marked pleurite and marked coxal margin in level F. At the posterior extremity of the pleurite and coxal rim there is no true articulation, figure 32G. The expanse of flexible pleuron is a little less, so that some support is given to a levator-depressor movement of the coxa on the horizontal pleurite. The coxa-pleurite junction is in effect a long hinge joint, but a posterior articulation is absent. The promotor-remotor movement of coxa2 and 3 takes place mainly between the pleurite and the body. The anterior end of the pleurite is firmly supported on the tergite (see above) and movement between this pleurite and the body take place at the freer posterior end.
62 264 S. M. MANTON The coxa-body articulation of leg 1 is rather different. This leg needs greater freedom of movement than legs2 and 3 because it is shorter, and needs to move through a larger angle in order to take the same stride, and it needs greater powers of promotor movement (figure 7a). Two pleurites X and Y (figures 31b, 32A) support coxa 1. They do not move on the body and are supported differently from the single meso- and metathoracic pleurites (see above). An elaborate coxal articulation with pleurite Y lies in the middle of the lateral proximal rim of the coxa. There is a deep and a superficial link of thin, highly sclerotized but flexible cuticle between the complex articulating facets, a condition found in essentials in the most strongly constructed of myriapodan joints (part 6, text-figs. 13, 15K). Coxal movements take place in all directions about this articulation, which does not restrict to the levator-depressor movements as on coxa 2 and 3. (iii) Leg movements in the Thysanura The leg action of the Thysanura differs from those of all other apterygotes in that the movements are best described as jumping, rather than walking or running. Unstable moments, with only two legs in contact with the ground, are usual, whether the legs of a pair are in similar phase, as in machilid jumping gaits, or in alternate phase, as in the Lepismatidae (8 6C). An essential feature of the jumping movement probably resides in each leg exerting a maximal thrust at the beginning of the backstroke, as in the bipedal spring of a jerboa, or at the moment when the leg of a galloping horse strikes the ground. In the horse a leg gains considerable momentum in a backward direction before it strikes the ground, and its leg is in contact with the ground for only part of the backstroke, the foot being raised before the end of this movement. The latter feature has been observed in Chilopoda running at their fastest* but not in Thysanura. The former has been demonstrated in Petrobius (8 6C(iii)) and may be expected to occur in Lepisma. The forward swing of the leg of Petrobius contrasts with other hexapods in that the movement is not executed evenly, as in the gait diagrams in figures 2-7, but is completed at great speed during the first part of the forward movement (8 6C(iii)). The rest period which usually follows is probably eliminated when the animal goes all out. As noted by Gray (1968), a limiting factor to the gaining of speed by galloping in mammals may be the ability to perform the recovery forward movements with sufficient rapidity. The habitually rapid forward swing of the leg in Petrobius may be a defence tactic which leaves the animal for ever at the ready to turn on its fastest jumping. The effectiveness of a horse s gallop resides in leg length, digitigrade foot, a wide angle of promotor-remotor swing, rapidity of backward acceleration of the leg before each footfall and rapidity of performance of the whole cycle. The leg is lightly built in the distal direction, with short proximal bones, proximal concentration of the muscle mass and suitably strong joints limiting movement to one plane, etc. (Howell, 1932, Gray, 1968). Most of these * Part 3, text-fig. llc shows foounarks curving towards the bottom of the page before sweeping upwards (forwards), these are made as the limb tip is raised. At slower progression, text-fig. lla, the marks sweep directly forwards, there being no backward swing after the leg leaves the ground.
63 EVOLUTlON OF HEXAPOD LOCOMOTORY MECHANISMS 265 locomotory and structural features are present in the Thysanura, elaborated in different manners by the Lepismatidae and Machilidae. The unique longitudinal hinge-like joint between coxa and pleurite on legs 2 and 3 (figure 31b) and the very strong arrangements on leg 1, provide a strong but supple base for adductor movements bringing the coxae together below the body before a jump and or the strong, rapid straightening of the leg at the jump, coxal abductor movements of small amplitude contributing to this. These movements are superimposed upon wide promotor-remotor action, taking place, not at the coxa body junction, but between pleurite and body on legs 2 and 3 ( 5 7C(i), (ii) above). The structure of the coxa-trochanter joint and its muscles (see below), are an integral part of this jumping mechanism, being suited to provide rapid, strong and precise movements in a manner not found in other hexapod classes. In the Lepismatidae there is no bipedal jumping, and no long horizontal coxa-pleurite hinge line is present as in Petrobius, because no comparable abductor-adductor movements are employed. The legs do not thrust the body upwards at right angles from the ground. Lepisma progresses by a series of rapid, one-legged hops, a most striking feature being the merging of the coxal promotor-remotor with the adductor-abductor movement. The coxa is very wide, housing massive muscles, it is almost horizontal in position with a small almost horizontal range of movements towards and away from the middle line, in the slot situated between the paratergal lobes and the sternite flanges (figure 34). The wide, strong and very rapid depressor-levator movements of the slender telopod thus become the stepping or hopping movement of the leg. The coxa-trochanter joint, is essentially as in the Machilidae, but modified remarkably so that the telopod can levate, not only up to the coxa, but postero-dorsal to it, as shown in figure 34. This most remarkable joint (see below) permits the short telopod to swing through a very large angle, enabling it to take a very long stride. But the movement is executed almost horizontally, there is no thrusting of the body upwards away from the substratum, as in Petrobius. These features assist Lepisma in gaining great speed from short, moving parts, in negotiating inclined or vertical surfaces and in penetrating small spaces without hinderance from projection by the legs. Such a leg, remarkable both in its movements and its morphology (see below), has not been found in any other class of hexapod or myriapod. Possibly some specialized pterygote has discovered the lepismatid solution to the same problems, but the generalised pterygotes have not. The lepismatid leg is a specialization of the type found in Petrobius, this type of coxa-trochanter joint being an essential pre-requisite to lepismatid evolution. (iv) Coxal musculature of the Thysanura The meso- and metathoracic pleurites of Petrobius as well as those of Lepisma move on the body with their coxae. Pleurite muscles, which are in effect extrinsic leg muscles, will be considered below along with muscles from the limb itself. Protractor and retractor muscles from the coxa. In Petrobius stout coxal protractors leave the strengthened anterior rim of the mesothoracic coxa (figure 32E, muscle 101 of Barlet (1967) for Lepismachilis) and pass forwards
64 266 S. M. MANTON and upwards to the tergite (figures 32D, C) while the protractors from the pleurite leave it as shown (muscles 113, 114 of Barlet), at a very poor leverage, as is so common in muscles causing a recovery, as opposed to a propulsive, movement. The pleurite protractors of Lepisma are seen leaving the anopleure and trochantin in figure 3 5a. Tiegs (1955) finds no coxal protractor muscles. It is probable that the protractor muscles, already partly arising from the coxa and partly from the pleurite in Petrobius (meso- and metathorax), have shifted to an entirely pleural origin in Lepisma, in association with the unusual axis of movement of the coxa on the body and the necessary mobility of the pleurites. Contraction of the tergo-pleurite muscles must pull the pleurites in towards the body, increasing their mutual overlaps, and this movement will also provide coxal promotion sufficient to contribute to the described movement and elongation of the relaxed coxal retractors. No such arrangement for coxal promotion has been found in any other class of hexapods. Plenty of examples have been given in Parts 7, 8 of muscles which have shifted their origins or insertions or both in the myriapods. The coxal retractor muscles of the mesothorax of Petrobius arise as stout sectors from the posterior and postero-mesial margin of the coxa, muscles 103, 104 (figure 32G) and pass upwards to the tergite, fanning also forwards (figure 32F, E, D). A pleural retractor (muscle 115, figure 32F, G) of large size leaves the flexible pleuron near the posterior limit of the pleurite and passes upwards to the tergite. These retractor muscles deform the flexible pleuron behind coxa and pleurite, pulling these structures inwards to the body, just as the emarginated coxa and the trochanter rim of Chilopoda are pulled in (Part 8, p. 306, figure 58). The metathoracic protractors and retractors resemble those of the mesothorax. In Lepisma the retractor muscles from the coxa insert upon the tergites much as in Petrobius and are not drawn here (see Barlet, 1954). Tiegs (1955) figures two stout tergal retractors from the coxa, t. cx. 3., t. cx. 4. and two sternal retractors s.cx.i., s.cx.2. The remotor movement of the coxa is thus well provided for, as is necessary for its strong and rapid movements. The prothoracic protractor and retractor muscles in Petrobius are differently contrived, and, since the pleurites here do not move with the coxa, there are no pleural sectors of these muscles. Stout protractor muscles leave the proximal anterior strengthened rim of coxa 1 and pass upwards and forwards to insert on the tergite and occipital sclerite (muscles 95, 96, Barlet, 1967). The retractor muscles 98, 99, 100 arise from a large coxal apodeme, projecting inwards and upwards from the coxal rim close to its articulation with the pleurite (figure 32A). These muscles slope forwards to the tergite, thus levering the coxa against its pleural articulation. The muscies do not clutter up the posterior margin of the coxa and thus space is left for the exit of muscle dep. tr.1. from the trochanter (see below). Abductor and adductor muscles from the coxa. These muscles on the mesoand metathorax of Petrobius cause the levator-depressor swing of the coxae on their horizontally placed pleurites which do not participate in this movement. The muscles are stouter and probably stronger than those in other apterygote classes (figures 32F, 35d). The rapidity of coxa abduction is probably important in providing some straightening of the leg at the jump, although the amplitude of the movement is not great (see above 6 7C(ii)). In Lepisma the
65 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 267 amplitude is also small (figure 34b), but is an essential component of the unique stepping movements. Coxal abductor muscles. In Petrobius abductor 102 leaves the pleuron close to the lateral coxal margin and passes upwards and forwards to the tergite above (figure 32D, C). Another abductor, muscle 136 of Barlet (1967), leaves the lateral coxal rim and passes inwards and upwards to the pleural apodeme (figure 32E, E). In Lepisma, one large strong abductor (muscle 135 of Barlet (1954) and muscle t.cx.5. of Tiegs (1955) in Ctenolepismas) leaves the dorsal coxal rim at its pleural articulation, as shown on figure 35d. There are some smaller muscles also (Barlet, 1954). The origin ensures that only abductor movement results. It is probable that the slot of thin, highly sclerotized cuticle near the dorsal rim of the coxa, shown in figure 35e, is of mechanical significance in easing the very rapid coxal abductor-adductor movements. The antagonistic coxal adductor muscles form two series, those roughly dorso-ventral in position, inserting on the tergite above, and those more ventral and horizontally placed. In Petrobius muscles 91, 92 leave the mesial coxal margin (figure 32D) and pass upwards and forwards to the tergite (figure 32C). In CtenoZepisma muscles t.cx.z. and t.cx.2. of Tiegs (1955, figure l), from the inner anterior and inner posterior rim of the coxa respectively, pass upwards to the tergite. In Petrobius one large ventral adductor, muscle 86, leaves the outer proximal face of the mesothoracic coxa and passes upwards, backwards and towards the middle line to insert upon the tendinous endoskeletal complex, a stout fan of tonofibrils supporting the endoskeleton at this point (figure 32F). There is no corresponding transverse adductor on the metathorax, because the coxa is more ventrally set on the body, and so there is no straight course open for such a muscle. In Lepisma the ventral adductors are more numerous, large, and as long as can be contrived in the space, features in keeping with the unique combination of adductor and remotor movements. Barlet (1954) describes muscles 106, 108 and 110. The origin of muscle 110, from the stout lateral marginal sclerotization of the coxa, is seen in figure 35e. Below it lies the origin of muscle 106 from a tendon, shown as a coxal process projecting as far as the word coxa on figure 33a, which is anchored in a coxal groove, cut in figure 3 5e. The groove is a strengthening device. Muscle 108 leaves the coxal face below the groove. All these muscles are stoutly supported by tonofibrils, which cannot be shown adequately in figure 35e. Muscles 106 inserts on the prothoracic endoskeletal plate, muscle 108 on the mesothoracic endoskeletal plate and muscle 110 inserts on the mesothoracic sternite near the middle line. The strength of the adductor muscles is correlated with the coxal movement shown in figure 34b leg 2, the swing towards the middle line assisting the backward movement of the telopod. The adductor and abductor muscles of the prothorax of Petrobius differ somewhat from those on the meso- and metathorax because of the difference in the coxa-body articulation and the presence of the coxal apodeme, which acts as a lever on the whole coxa. Muscle 97 of Barlet (1967) would appear to be an abductor, resembling abductor 135 of Lepisma (figure 35d) in arising close to the coxal articulation. Muscle 97 may also be a stabilizer. There are other prothoracic muscles which appear to be stabilizers, rather than principally causing movements, such as muscle 110 of Lepisma (Barlet). The prothoracic adductors correspond with those of the mesothorax. The transverse
66 268 S. M. MANTON adductor 81 (figure 32A) corresponds with muscle 86 (figure 32F) and dorsal adductors 89, 90 (figure 32A) pass from the mesial coxal margin and face to the tergite above, as do muscles 91, 92 on the mesothorax (figure 32D, C). Other coxal muscles. As in many animals with complex musculature, a precise function cannot be ascribed to every muscle because their actions depend upon the tension of a number of other muscles. The coxae of Lepismachilis and of Lepisma (Barlet, 1967, 1954) bear small muscles which insert on the tendinous endoskeletal plates. Lepisma also shows muscles 107, 109, 11 1 from the coxa inserting on the preceding endoskeletal plate, the sternite and the endoskeletal plate of the same segment (mesothorax in Barlet, 1945, Figure l), These muscles, by their wide spread, probably stabilize the coxa and contribute to the remotor movement. (v) Leg structure and the special proficiencies of the Thysanura The leg movements used in the jumping gaits of Petrobius, so unusual for a hexapod, and the peculiar stepping by the telopod in Lepisma have been described above (0 7C(iii)). The coxa-trochanter joint, one of the most remarkable among hexapods, provides rapid, very strong and wide levator-depressor movements, which are essential features of the proficiencies. With their evolution are associated the strong extrinsic trochanter muscles and the pleural apodeme used for their insertions. Morphology and movements of the coxa-trochanter joint. This joint is shown in figures 31, 36. The pivot articulations on either side between the two podomeres are not equatorial (half way between the dorsal and ventral faces of the leg) but lie close to the ventral face, at the base of an enormous apodeme from the trochanter which projects into the coxa and gives origin to intrinsic and extrinsic depressor muscles of great size. A normal position for the anterior pivot articulation is shown in Gzmpodea and Japyx figures 12F and 9a mesothorax, unlabelled. Similar positions are also seen in Lithobius, (Part 6, figure lla, Part 8, figure 58), Polydesmus (Part 6, text-fig. 12) and in the grasshopper (Snodgrass, 1929, Fig. 36A "f"). The leverage of the depressor (extensor) muscles in Thysanura lies between the tip of their apodeme of origin and the pivot articulation at the base of this apodeme. Doubtless this is a mechanically stronger arrangement than the usual one in myriapods and other apterygotes where the depressor (extensor) muscles pull from the ventral or mesial rim of the trochanter, moving it on an equatorial set of pivots, situated either on both faces of the leg, or strongly constructed on the anterior face only. Many Pterygota possess an apodemal origin of the trochanter depressors, as in the Thysanura (cf. Evans, 1961, Text-fig. 33, Snodgrass, 1929, Fig. 36A 71 Ap.). The most significant feature of the coxa-trochanter joint of Petrobius is the great range of movement which can take place at the proximal lateral (dorsal) rim of the trochanter. Here there is much emargination and the rim does not bear levator muscles, as is usual in other classes. Were such muscles present in Petrobius they would, at times, have to pull at very different angles from their tonofibrils, a condition which arthropods avoid, and the necessary changes in length of levator muscles so situated would be greater than they actually are. A more mechanically serviceable arrangement lies in the pillar-like swinging
67 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 269 tendon attached to the ventral trochanter cuticle a little distance from the apodeme, and from this tendon the levator muscles extend into the coxa. The tendon can swing on its fan of tonofibrils so that its levator muscles always pull in line with it, and not at an angle as drawn in figure 3 la. The muscles on the arcuate sclerite of the spider femur-patella joint use a comparable device, whereby they pull at a more or less constant angle on the sclerite, although the angular changes at the joint are very great (Manton, 1958a, Fig. 3). The collembolan trochanter tendon, in much the same situation as in Thysanura (figures 18% 23E, 24G, 29a, 30), is used largely by suspensory muscles preventing joint dislocation ( 8 7B(iv)). The levator muscles arise largely in the normal position, from the rim of the trochanter, but spread also on to the tendon. In Lepisma the coxa-trochanter joint has further elaborations, particularly on leg 1, enabling the levator-depressor swing of the telopod to take place over fields of movement such its in figure 7a for Petrobius, yet the coxa always remains backwardly directed as in figure 34. There is a twist in the hard parts so that the trochanter fits into a coxal depression on the postero-dorsal side, particularly marked on leg 1, thus the trochanter carries the telopod clear of the coxa when levator movements bring the telopod to an extreme forward position, with the femur situated between the coxa and the body, a unique leg movement by a unique joint. Distally the trochanter is emarginated at its dorsal rim (figures 31, 36) so that the dorsal face is less than half the length of the ventral face in Petrobius and becomes almost non-existent in Lepisma. These features enable the enormous range of swing to take place, as shown in figure 36a for leg 2 of Lepisma, and a smaller range in Petrobius, but enough to bring the trochanter-femur both horizontal and a little upward in direction when the coxae are fully adducted below the body, an essential preliminary bending of the leg before the jumping extension. MuscEes of the coxu-trochanter joint. Muscles from the trochanter, both intrinsic and extrinsic, are massive and differ in important features related to their function from those of other apterygotes. The intrinsic levator and depressor muscles lev.& and dep.tr. fan out over the antero-lateral and postero-mesial faces of the coxa respectively, inserting mainly near the proximal coxal rim, so that maximum length is obtained (figures 31, 36). The great width of the coxa in Lepisma houses a particularly large bulk of these muscles which contribute to the very strong and rapid stepping by the telopod, the levator-depressor movements having become in effect the stepping action of the leg. The extrinsic depressor muscles dep.tr.2. from the trochanter in both Petrobius and Lepisma are large and long. They separate from the intrinsic depressors within the coxa. In Petrobius they form three closely applied sectors (muscles 127, 128, 129 of Barlet, 1967), which insert all round the pleural apodeme (figures 3 1, 32D, E, F). On the prothorax, in the absence of a pleural apodeme, these muscles insert on the tergite and endoskeletal plate (123, 125, 126, Barlet, 1967). An exit space from the coxa for muscles 125, 126 is made by the absence of coxal retractors at the posterior coxal margin (see above). In Lepisma the long depressor muscles leave the apodeme on the trochanter as in Petrobius, they do not insert on the pleural apodeme
68 270 S. M. MANTON but on to the trochantin (muscles 122, 123) and tergite (muscle 124, all of Barlet, 1954), muscle 124 being the largest. The great width of both body and coxa places the trochanter apodeme nearer to the middle line of the body than to the pleural apodeme above, and the dep. tr.1. muscles pass directly upwards. Extrinsic depressor muscles from the trochanter are present in all myriapod and hexapod classes, and, with the intrinsic depressors, provide a major part of the propulsive thrust by the leg. But the extrinsic depressors of the Thysanura are much more robust and bulky than are those of other apterygotes (cf. Campodea, figure 12F dep. tr.2., Tomocerus, figures 22-24, dep. tr. I.), suggesting that they can exert considerable force. The large displacements of the trochanter at its proximal joint can be done with minimal length changes of the muscles, because the leverage between the tip of the apodeme and the articulation is not great, a feature also favouring high frequency of stepping (short pace durations) of Lepisma. Extrinsic levator muscles from the trochanter. These are of common occurrence in myriapods where there are usually no coxal levator movements. The raising of the trochanter lifts the whole telopod (Part 8, figures 53b, 63b; Part 9, figure 5e). Extrinsic levator muscles from the trochanter are well developed in Lepisma but not in Petrobius or in other apterygote classes. These muscles separate from the intrinsic levators and pass directly upwards in two groups of sectors, lev. tr.1. ka. inserting on the pleural apodeme, figure 3 5d, e, (muscle 193 of Barlet) and 1ev.tr.l.an. inserting on the anopleure, figure 35c, d, e (muscle 195 of Barlet). The presence of strong extrinsic levator and depressor muscles from the trochanter of Lepisma is correlated with the very rapid and wide stepping by the telopod, the coxal movements being small. Both the strong and supple coxa-trochanter joint and its very strong musculature in Lepisma are more specialised than in Petrobius and provide part of the means whereby the adductor-abductor movements become the effective stepping by the leg. These short legs take a remarkably long stride (8 6C(v)) yet project but little beyond the flanks. In no other hexapod or myriapod has a comparable differentiation in the range of movement along the leg been found. This differentiation is a most remarkable thing in itself. The coxa-trochanter joint and its muscles in Petrobius are suited to give sudden and maximal leg extension, as is needed for jumping, in contrast to the walking and running movements during which the thrust on the ground is roughly constant during the backstroke. But the morphology and achievements of Lepisma need a coxa-trochanter joint and muscles much as in Petrobius as a starting point for their further advances. This part of the thysanuran leg thus becomes one of the most important features of their organization in contrast to other hexapod classes, and indicates an ancestral long standing tendency towards jumping as opposed to walking movements. Muscles and joints of the telopod There is little movement at the trochanter-femur joint in either Petrobius or Lepisma The joint is stabilised by a retractor femoris muscle, ret.fe., as in many other arthropods (Part 6, text-fig. 18h, j; Part 8, figures 47 ret.p. fe., 54 ret.fe. etc.). The femur-tibia pivot joint is very mobile, the flexor muscles Fti. being remarkably bulky in Lepisma, giving strength to the stepping by the slender distal podomeres, and long in Petrobius, extending forwards into the trochanter, as suits the
69 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 271 production of large displacements at the joints used in jumping. The dorsal hinge joint between tibia and tarsus is particularly strongly made in Petrobius. The flexors to the tarsus jl.ta. are similar in Petrobius and Lepisma, but the flexors to the tarsal claws are much longer in Petrobius, some sectors extending to the anterior part of the long femur. Thus leg structure in Thysanura has common features which are characteristic of this class in contrast to the legs of other classes, but modified towards the very different leg action and needs of the Machilidae and Lepismatidae. (vi) Summary of thysanuran peculiarities of thoracic and leg structure in association with habits Since so much has been written concerning the primitiveness of the Lepismatidae, or of parts of their morphology, and of the possible ancestral connection between the Thysanura and Pterygota, it seems fitting here to list the specializations of the thysanuran thorax, described above on a functional basis, which show great divergencies from other hexapod classes. A dichotomy from a type of animal inclined towards jumping has apparently resulted in the contrasting features of the Lepismatidae and Machilidae. The former practise a unique leg action and the latter have perfected jumping gaits. The pleural armature in both gives strong coxa-body support by mobile pleurites, whose mobility is quite unlike that of myriapods or any other class. No other hexapod or myriapod shows a promotor-remotor leg movement sited at the pleurite-body junction, as on legs 2 and 3 of the Machilidae (with differences on leg l), while the adductor-abductor movement situated between coxa and pleurite on a long hinge is not seen elsewhere. Only the Lepismatidae among known hexapods combine the promotor-remotor swing of the leg with adductor-abductor movements, so that stepping is largely done by the levator-depressor movement of the short, fast moving telopod. Only the Lepismatidae show a shift in origin of all the coxal promotors to the pleurites, whose movements promote the coxa. Both orders need the remarkable coxa-trochanter joint, with the unique excentric articulating facets at the base of a huge apodeme used by depressor muscles, and a swinging tendon for the origin of the levator muscles, of the trochanter. Extrinsic trochanter muscles are more fully developed than in other hexapod classes. These features enable a maximal thrust against the ground to be exerted at the beginning of the backstroke, giving the jumping movements. The further development of the coxa-trochanter joint in the Lepismatidae, so that the levator movement of the telopod brings it not only up to the morphologically dorsal face of the coxa, but further up and alongside it, gives the enormous range of swing of the lightly built distal part of the leg. Only in the Lepismatidae, among known hexapods, is there a differentiation in the angle of swing of the leg into the strong movements of small amplitude by the coxa and the extraordinarily wide range by the telopod, whose swift movements have defied a recording of their maximum frequency. The Lepismatidae also achieve a body shape capable of very fast darting running, with very little leg projection, thus favouring running in exposed places or under cover. Not one of these features can be regarded as a primitive hexapod
70 272 S. M. MANTON characteristic. The contrast with other hexapod classes is great; and advanced and efficient as are the legs of the Machilidae, it is those of the Lepismatidae which show the greatest specializations in correlation with their habits. 7D. THE PLEURON, LEG, LEG-BASE AND ASSOCIATED MOVEMENTS IN THE PROTURA The general form of the thoracic segments in the Protura (figure 9b, c) resembles those of the Chilopoda and Diplura. In these animals the basic structure provides two assets, namely trunk flexibility and strong support for the coxae (5 7A). Well formed tergites and sternites are separated by a wide expanse of flexible pleuron bearing a number of pleurites (figure 9a and Part 8, figures 48,49, 58). These general similarities are associated with roughly similar ways of life and methods of gaining shelter. The coxae of legs 2 and 3 of the Protura are laterally directed and deeply set in the lateral margin of the sternite, leaving a narrow, well sclerotized sternal bridge between each pair of coxae (figure 9c), as in the Campodeidae (figure 12a, c, d). The prothorax shows differences due to the position and use of the prothoracic limbs. The Collembola and Thysanura Machilidae contrast in possessing no thoracic sternites, their pleuron is restricted, they have few pleurites and their coxae are pendant and ventrally directed (figures 23, 24, 31, 32). The legs of Protura and Diplura Japygidae are short, strongly constructed and used in a similar manner. Owing to the greater width of the abdomen than the thorax, leg projection is minimal, so giving least hindrance when the animals are moving in the soil (pls. 1, figures i, j; 2, figure h). A rocking mechanism of the leg, which assists the propulsive extension of distal hinge joints lacking extensor muscles, is present in Chilopoda, Symphyla and Pauropoda (Part 8, figures 2, 3; Part 9, pp ). The Protura also possess a rocking mechanism to the coxa, but quite differently contrived from that of the myriapods, and differing also from the unique rock in the Diplura situated in the leg itself. (i) Thoracic flexibility of the Protura The whole body of a proturan is very flexible both in bending about the longitudinal axis and in extending and shortening the thoracic as well as abdominal segments. Thoracic flexibility depends upon narrow intersegments and a flexible pleuron which will bend between the pleurites, as in aiplura, but the correlated tergite and sternite flexibility is differently contriv=d. The proturan tergites are well sclerotized and large anteriorly. They show a transverse break in the sclerotization at the anterior level of the coxae. The sternites are most solidly sclerotized between and behind the coxae. Just anterior to the coxa-sternite articulation there is a transverse band of lesser sternal sclerotization. These two sites of flexibility are marked by arrows on figure 9b, c, and with the flexible pleuron, make possible easy outward bowing of the flanks and dorsal and ventral surfaces when the musculature shortens the thoracic segments. There is no detailed similarity between the pleurites of. Diplura and Protura (figure 9a, b).
, shown in figure 9b as pleurite 1.")
71 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 273 (ii) The coxa-body articulation and its movements in the Protura As shown by Prell (191 3) the proturan coxa has two articulations, ventrally with the sternite and dorsally with the middle of a large arc-shaped pleurite (the trochantin or subcoxal segment of entomologists), shown in figure 9b as pleurite 1. The axis of the protractor-retractor swing of the coxa is thus approximately dorso-ventral and restricted to the transverse plane, as in Diplura, where only a coxa-sternite articulation is present (figure 9a). In no other apterygote class are there two such coxal articulations with the body. This provision must strengthen the movements of the stout proturan legs against soil and other particles among which the animals live. The ventral coxa-sternite articulation on the mesothorax receives no particular support. Berlese (1910) and Prell (1913) shows a median apodematous ridge lying longitudinally on the meso- and metasternite of the mesothorax. The ridge reaches the posterior extremity of the metasternite and extends anteriorly across the zone of lesser sclerotization to reach the posterior border of Prell s presternite (figure 9c). This ridge bears the insertions of the coxal protractor and retractor muscles, figured by Berlese (1910), while the extrinsic depressor muscle from each trochanter crosses over to insert on the sternite margin of the opposite side. There are also a pair of stout protractor and retractor muscles from the coxa to the anterior (most solid) part of the tergite. On the metathorax the apodematous sternal ridge figured by Prell (1913) forks anteriorly to end at the coxa-sternite articulation (figure Sc), roughly resembling the supporting arms of the Y-skeleton of the Diplura which articulates with a pair of coxae on every thoracic segment (8 8A). Posteriorly Prell shows the metathoracic apodematous ridge to fork and pass directly laterally to the sternite margin opposite the small metapleurites. These arms bear the retractor muscles from the coxa. These meso- and metathoracic apodemes show only a limited resemblance to the Y-skeleton of the Diplura. Nothing closely resembling the dipluran spine-apodeme is present. This hollow structure projects inward and posteriorly from the posterior part of each thoracic sternite (figure lo), and carries the huge coxal retractor muscles 6 and some ventral longitudinals. There are no apodemes on the prothoracic sternite of Protura. (iiil The extrinsic rocking mechanism of proturun legs and leg structure Superimposed upon the simple promotor-remotor swing of the coxa and leg is a rocking movement uniquely contrived. Pleurite 1 (the subcoxa or trochantin of Prell) does not touch either the coxa or the sternite, except at the dorsal articulation. But pleurite 2 (the mesopleurite of Prell), which forms another extensive arc beyond pleurite 1, unites with the sternite in front of the coxa and is thereby futed in position. In the middle of the ventral border of pleurite 2 is a large pair of sliding facets placed horizontally between the pleurites (figure 9b). From this articulation pleurite 2 is prolonged into an upward and anteriorly directed arm, with an internal apodematous ridge bearing muscle insertions (Prell, 191 3). Photographs of walking proturans show that a rocking of legs 2 and 3 takes place during stepping, as in Chilopoda, Symphyla and Pauropoda. During the backstroke the dorsal face of the leg is
72 274 S. M. MANTON rocked anteriorly, and the reverse occurs on the recovery forward stroke (see 8 2). This movement must be implemented via the sliding facets between pleurites 1 and 2, pleurite 1 with the appended dorsal coxal articulation, being pulled forwards relative to pleurite 2, by muscles inserted on the apodeme of pleurite 2. The walking legs 2 and 3 of the Protura possess only one pivot joint with an antagonistic pair of muscles, that between coxa and trochanter. The distal trochanter joint moves but little and is followed by three hinge joints with flexor, but no extensor muscles (Prell, 1913). This is a longer series of extensorless joints than in any other class of hexapod, the Diplura, Collembola and Thysanura possessing pivot joints, or modifications thereof, as far as the tibia (figures 9a, 12a, 30, 36). The effect of this rocking of the coxa is similar to those of the differently contrived rocking in myriapods and Diplura. In all the dorsal leg surface and its hinge joints are brought a little anterior in position during the backstroke, so assisting the extension of flexorless joints during the propulsive backstroke and easing the work of the long and strong proturan depressor muscles from the trochanter. 7E. CONCLUSIONS CONCERNING THE PLEURON AND LIMBS OF THE APTERYGOTE CLASSES It has been shown above how very different are the Diplura, Collembola, Thysanura and Protura in their external pleural morphology and cox4 articulations, and how the peculiarities of each class serve functional needs. The differences between the podomeres, their jointing and musculature in these classes are also striking and have been related to function. The class differences are such that preconceived schemes of basic pleurite and limb features, devoid of any functional considerations, become untenable. (il The pleuron In the Diplura the pleuron serves the needs of trunk flexibility; it is wide and exposed and is strengthened by pleurites. The same is true of the Protura, but the pattern of pleurites is different (figure 9a, b). The Collembola and Thysanura contrast in possessing a rigid thorax, the pleuron is smaller in extent, covered by paratergal lobes etc., and possesses fewer and smalier pleurites. In the Chilopoda (Part 8) the extraordinary flexibility of the fully armoured body of the Geophilomorpha was related to shape changes during burrowing and the delivery of a thrust against the soil, in contrast to the less sclerotized pleuron of the other three chilopodan orders possessing fewer and smaller pieurites. While this remains true, comparison with the apterygotes suggests that it is the need for trunk flexibility which leads to good pleurite development in both Geophilomorpha, Protura and Diplura. In the more rigid bodies of anamorphic Chilopoda, Collembola and Thysanura the pleurites are smaller and fewer. The thoracic flexibility of the Protura, provided by lines and tracts of lesser sclerotization across tergites and sternites and by the wide pleuron, shows no similarity in detail with the corresponding parts of Chilopoda and Diplura. The great lateral flexibility of japygids is provided by the many vertical lines of flexure between pleurites, pre- and intercalary sternites, which invade the pleuron (the campodeids are less fully sclerotized and less flexible). The pro-
73 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 275 and metacoxa of Chilopoda are represented in position by fixed expansions of the dipluran sternite, supporting the axis of swing of the leg. Mobile pro- and metacoxae are not required. Dipluran pleurites lying in the areas labelled trochantin and distal anapleurite by Barlet & Carpentier (1962) and subcoxa 1 and 2 by Denis (1949), and trochantin (pleurite 1) and pleurite 2 here are only roughly comparable with the katapleure and anapleure of Chilopoda. They support the fixed axis of coxal protractor-retractor swing in the Diplura and do not provide the rocking movement of the coxa exhibited by so many myriapods. Proturan pleurites provide a rocking mechanism, but in an unique manner. Pleural armature is very various on the different segments of some species in both Chilopoda and Thysanura, and the form and number of pleurites is associated with functional needs in the several apterygote classes. The collembolan pleuron is much restricted because of the hydrostatic mechanism working the jumping organ, the tergites spreading laterally over the leg-bases (cf. Diplura). Only one pleurite is present (metathoracic) and there is ample provision against hydrostatic bulging of the rest of the pleuron. The single pleurite corresponds with the katapleure of Carpentier (1949) but its correct shape has not been recorded before. The anapleure of Carpentier on leg 3 is shown to be the pleuron and not a sclerite. The zones marked trochantin on legs 1, 2 and 3 by Carpentier are shown here to be the undifferentiated pleuron on the prothorax and the proximal coxal segment of legs 2 and 3 respectively. The bulges above leg 1, the uochantin of Carpentier and subcoxae 1 and 2 of Borner (1921), and simply the pleuron, bulging between the endoskeletal ties which hold the pleuron firmly in place against hydrostatic dilatation. Evidence has been presented to show that the complete division of coxa 2 and partial division of coxa 3 is a functional service associated with the unique suspension of coxae from the body and the required movements. There is no sound evidence suggesting that the proximal parts of coxae 2 and 3 are formed by a metamorphosed pleurite, the trochantin. The pleuron of the Thysanura also is restricted in extent and well boxed in by the tergite for functional reasons. The number and shape of the pleurites also serve the animal s needs. One well differentiated pleurite on meso- and metathorax of Petrobius, two on the prothorax of Petrobius and three on the meso- and metathorax of Lepisma all serve particular needs, and there seems no reason why these pleurites should be forced into a common scheme of terminology. There are plenty of examples among Chilopoda showing different numbers of pleurites on consecutive segments; there is no reason why the single pleurite of the Symphyla should be regarded as the homologue of any one chilopodan pleurite. The three pleurites in Lepisma termed trochantin, katapleure and anapleure by Barlet (1954), are all true pleurites in that they are strongly sclerotized areas. But the machilid interpretations by Barlet ( 1967) cannot be upheld. The two pleurites on the prothorax he regards as anapleural, the flexible pleuron, a most necessary area between the pleurite and tergite on meso- and metathorax, is called a membranous anapleurite, here considered to be a contradiction in terms. The overriding desire to find homologous structures everywhere has led to the identification of the strengthened proximal margin of the coxa of Petrobius, marked coxal margin on figure 34D, F and drawn on a larger scale in E, as a trochantin (Barlet, 1950, 1967). This thickened, folded coxal margin
74 276 S. M. MANTON braces the coxa against the pull in many directions of extrinsic and intrinsic muscles. The thickening does not extend to the mesial side, where it is advantageous for the retractor muscles to pull in the postero-mesial margin of the coxa, deforming the pleuro-sternal cuticle. In Lepisma there is a corresponding and elaborate coxal margin near the coxal articulation b, and there is a trochantinal pleurite in addition. There are many examples among the myriapods of thickenings and foldings at the margins of coxa, trochanter, diplosegmental tergites and sternites, etc. (Parts 8, figures 46 skeletal annulus of trochanter, 53 similarly drawn but unlabelled, Part 7, text-figs. 7 see unlabelled prophragma, 17, 18 marked prophragma, etc.). The identification of part of the margin of the coxa of the Machilidae as a trochantin cannot be entertained. The presence in Lepisma of three vertical pleurites anterior to the coxal base is associated with the most unusual position of the coxa and the partial fusion of the protractor-retractor and abductor-adductor movements. Retractor coxal movement is eased by a sliding apart of the overlap of the pleurites, so allowing the coxal rim to ease away from the body anteriorly while its posterior rim is pulled in. It is suggested here that the variety in pleurites found in the several apterygote classes and even in families of the same class, or on different thoracic segments in one species, cannot be forced into a common scheme. On the contrary they all represent different solutions of the various functional needs of the animals. In Part 9, figure 10a the locust was taken as a pterygote example for comparison with the myriapod classes. A major pleurite in the locust, united to both tergite and sternite provides the principal coxal articulation and support. The trochantin, situated in the arthrodial membrane at the leg base anterior to the main coxal articulation, is moveable and provides a secondary articulation with the coxa on some segments only in some insects. The so called trochantin of entomologists is a true pleurite in Lepisma and in the Diplura. But this trochantin forms the principal and only coxal articulation in Lepisma, and is not articulated with the coxa in the Diplura. Comparison between the locust and Lepisma (Part 9, figure 10a and figures 33, 35a, c here) might suggest that if any homologies exist between their pleurites, those named trochantin do not correspond, the principal pleural articulation of the locust being the probable homologue of the articulation between coxa and trochantin of Lepisma. The view that pleurites originally formed concentric rings round the base of the coxa (Matsuda, 1963) has nothing to commend it because there would be no apparent function for such structures and the necessary coxal articulation with the body is not catered for. Pleurites 1 and 2 of the Protura are the most arc-shaped pleurites among the hexapods, but this shape implements the proturan coxal rocking mechanism which is so differently contrived from the rocking in all other classes. It is considered here that none of the described structures in or near the base of the leg in any apterygote can be regarded as the remains of a subcoxal leg segment. (ii) The leg and leg-base The contrasting coxal body unions and coxal support of the Diplura (sternal), Thysanura (tergal via a mobile pleurite) and Collembola (suspensory
75 EVOLUlION OF HEXAPOD LOCOMOTORY MECHANISMS 277 system from the endoskeletal tendinous plate) appear to be mutually exclusive and the result of independent evolution. In all three classes extrinsic leg muscles utilize a site for insertions situated internally in the body, besides the surface sclerites. The Diplura employ a median sternal spodeme, differently arranged in Campodeidae and Japygidae (6 8), the Collembola utilize an endoskeletal node of connective tissue situated over the mouth of the coxa (5 7B), and the Thysanura possess a pleural apodeme serving the same purpose (see above). Again, these sites for extrinsic muscle support are so different as to imply independent evolution. Chilopoda use the segmental tendons in the longitudinal musculature; the Symphyla use a coxal apodeme; the Diplopoda a trachael pouch apodeme; all are different solutions of the same mechanical need. The Protura contrast with the other three apterygote classes in possessing both a sternal and a pleural articulation to the coxa. The extrinsic leg muscles are chilopod-like in that they utilized the tergal and sternal surfaces for their insertions. The only cuticular elaborations for muscle attachments are simple ridges on sternites and pleurite 2 (Prell, 1913). There are no long hollow apodem es. The myriapodan rock of the coxa on the body, additional to the normal promotor-remotor swing, which eases leg extension in the propulsive phase, is present only in the Protura, but is differently contrived. The Diplura have a unique rock within the leg itself serving the same need. Levator-depressor (abductor-adductor) movements at the coxa-body joint are largely absent in myriapods and are not possible to Protura. Coxal levator muscles occur in the Diplura and depressor muscles also in Cumpodea but not Japyx. Levator and depressor muscles from the coxa are very well developed in Collembola and Thysanura. Thus there is no similarity between the leg-base of any apterygote class and any myriapod. The limb podomeres and musculatures of the apterygote classes show many specializations not found in myriapods, such as all the features associated with the extreme flexibility of the coxa-trochanter joint in Collembolan and Thysanura; the unique suspensory system of collembolan legs and their series of tendons and muscles which prevent dislocation at the joints under internal hydrostatic pressure at a jump; the small number of podomeres in the telopod of Collembola, etc. All these features, together with those of the pleuron just considered, point to differences between the apterygote classes which are so fundamental as to indicate that they must have arisen and evolved in parallel and in no way one from another. Even the Thysanura show a line of thoracic evolution markedly divergent from that of the Pterygota. 8. THORACIC ENDOSKELETON AND MUSCULATURE Both these systems serve the differing thoracic needs in the several classes of hexapods. The characteristics of the musculature partly depends upon the nature of the exo- and endoskeleton. The latter has been described in accurate detail for a number of hexapods. But the authors of some of these descriptions believe that endoskeletal structures are homologous from class to class, although the component parts are made by apodemes, or by tendons, or by muscles in the various examples. It is considered here that such findings merely demonstrate a common functional need, which has been met by different
76 278 S. M. MANTON means in the several classes or smaller taxa. In short we have here a clear case of convergent and independent evolution of endoskeletal structures, not one of homology. Endoskeletal structures are very stable in type, but not in detail, within each class throughout the Arthropoda, and have been evolved many times (Manton, 1964). The trunk musculature and endoskeleton of the Diplura and Protura is of particular interest, in the light of work on these and other classes, because the details can be interpreted ro a considerable extent both functionally and phylogenetically. The thorax of Diplura and Protura possess a wide pleuron, many pleurites, well developed tergites and sternites, often subdivided, and laterally projecting coxae articulated with the sternites. These external features are also present in Chilopoda and Symphyla but not in Diplopoda or in the other hexapods. The exo- and endoskeleton and muscles of the former classes are associated primarily with thoracic flexibility in contrast to the Collembola, Thysanura and Pterygota where thoracic rigidity is required. Diplopoda also need rigidity, although they possess highly specialized mobility in the dorso-ventral plane. The morphology of Protura and Diplura associated with thoracic mobility does not appear to be basically primitive. It is alternative to the structural features giving rigidity of thorax in the other hexapods which must have been established early in their evolution. The thoracic endoskeleton and musculature of the Diplura and Protura is considered below. Those of the Collembola are described along with the abdomen because of the association with the jumping mechanism (8 9a(iv), (v)). The Thysanuran eridoskeleton and musculature has been described by Barlet, etc., references to this work being given in 8 lod(i). 8A. ENDOSKELETON OF THE DIPLURA AND PROTURA The dipluran endoskeleton is partly apodemd and partly tendinous. The details differ in the Campodeidae and Japygidae in association with long legs and fast running of the Campodeidae and strength for the leg-base of the J apygidae. The apodemes directly (Japyx) or indirectly (Chrnpodeu) support the coxa-body articulation (8 7A): they provide strong sites for insertion of coxal retractor muscles and the system promotes thoracic flexibility in a manner not found in myriapods. The simple apodematous ridges on the sternites of the Protura have been referred to above, along with the leg-base (8 7D(ii), and see figure 9c). (i) Japygidae General views of the ventral thoracic cuticle of japygids are given by Denis (1949, Fig. 47B) and by Barlet & Carpentier (1962, Figs. 1-3). On each thoracic sternite a Y-shaped ridge of cuticle is infolded, leaving surface grooves (figure 9a). The tips of the arms articulate with the coxae (5 7A(i)), and from the posterior end of the stem of the Y a stout hollow apodeme projects backwards into the following segment as the spina (figures 10, 1 le), bearing sectors of the ventral longitudinal and coxal extrinsic muscular systems (vzm.2., ret.co. 6. ). The length of the spina differs among japygid genera; in the small Solomon Island japygids it is relatively longer than in Heteroiapyx, and projects far into the succeeding segment.
.")
77 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 279 On the prothoracic sternite another smaller Y-shaped sclerotization lies anteriorly, the significance of which hitherto has not been understood. Its arms and stem provide sites for the origin of dorso-ventral muscles to the head. The skeleton is cut away in figure 10 anterior to the coxal level, but the muscles are drawn (unlabelled). Thus the extra Y-apodeme facilitates the marked flexibility of the head on the trunk noted above ( 5 6A(i)). A pair of endoskeletal tendons pass inwards and upwards from the arms of the Y-shaped skeleton well behind the level of the coxa (black on figures 10, 11F). These tendons bear an abundance of dorso-ventral muscles, inserting on their own and on the following segment, (dv., dv.i., dv.2., dv.3.) and sectors of the ventral longitudinal system (vim. I., vlm.5, vlm. 6., v2m. 7. ). (ii) Campodeidae The apodemal endoskeleton is more deeply sunk in the body and its origin hardly shows on the sternite surface. A median infolding from the posterior part of the sternite (figures 12b, 13a) corresponds with the stem of the Y in Japyx. The apodeme so formed expands above the nerve cord into a horizontal median endoskeletal plate corresponding with the spina of Japyx, but it does not extend back into the following segment. The median plate is supported anteriorly by pillars of cuticle from the sternite (figure 12FF) on the mesothorax and by pillars to the lateral arms of the plate lateral endoskeletal bars on the prothorax (figure 12E). Presumably these internal lateral endoskeletal bars correspond with the superficial arms of the Y-skeleton of Japyx. A deep dorso-ventral muscle dv. arises from the lateral edge of the median plate of Campodea (figures 12F, 13a), and very simple ventral longitudinal muscles extend from the internal tendons, anchored at the presternite bulges, to the lateral endoskeletal bars behind, and then on to the next corresponding tendon (figure 13a). (iii) Conclusions concerning the endoskeleton of the Diplura and Protura The evolution of the japygid spine apodeme thus appears to be associated with the marked strength of leg movements and with body flexibility. The central position of the spina enables the strong retractor leg movements to be uninfluenced by conspicuous local flexures of the thorax. Since the spina and its covering of muscles occupies so much of the middle body space, the formation of separate lateral endoskeletal tendons is a mechanical necessity. The more deeply set apodemal system of Cirmpodea, on the other hand, is associated with the longer legs and large coxae, the latter are deeply sunk into the sternite laterally, otherwise there might be unsuitable lateral projection, and between the coxae the sternite is narrow and much less robust than in Japyx. The proturan sternal apodemal ridges on meso- and metathorax differ on the two segments and show no exact resemblances to the japygid Y-skeleton. The essential functions of the Y-skeleton lie at either end, in the coxal articulation anteriorly and in the spina and its long muscles posteriorly. The rest of the apodeme is not clothed with muscles. In the Protura no sternal apodeme is required on the prothorax owing to the forward direction of the coxae. Their
.")
78 280 S. M. MANTON protractor and retractor muscles converge onto the posterior margin of the sternite, not to the middle line as on the meso- and metathorax (Berlese, 1910). The proturan sternal apodemes are best developed where the coxal protractor and retractor muscles from either side converge (Berlese, 1910, P1. XI, fig. 119 muscles spa,spp.). In some genera there is a mesothoracic internal arch of cuticle in others the apodeme is not sunk (Berlese, 1910, fig. 1, P1. 10, fig, 114). Only on the metathorax is there slight apodemal support of the coxa-sternite junction by the anterior forking of the apodeme drawn by Prell (1913, P1. 111). The apodemal sites for insertion of meso- and metathoracic coxal muscles are stronger than the simple flat sternites of Chilopoda, as is suitable for the fewer, strong proturan legs. There are no skeletal provisions for great leverage by the muscles, as in japygids. The resemblances between dipluran and proturan endoskeleton are convergent and inexact, as are the exoskeletal features permitting thoracic flexibility in the two classes ( 5 7D(i)). 8B. DIPLURAN AND PROTURAN MUSCULATURE AND ITS FUNCTIONAL ASSETS In both classes dorsal and ventral longitudinal muscles are present, but no lateral longitudinals, as occur in Chilopoda and Diplopoda. Antagonistic deep dorso-ventral and deep oblique muscles are vaiiously developed in Campodeidae and Japygidae, together with superficial muscles concerned with causing thoracic flexibility and endoskeletal muscles serving the same purpose. The Protura show muscles, more complex in arrangement than those of Campodeidae, which do not fall into the categories described for other hexapods. (i) Dorsal longitudinal muscles The Diplura possess parallel muscles extending from one tergite to the next (figures 10, 13a). There are no long muscles crossing more than one intersegment, as in Chilopoda, where it is essential to maintain body rigidity during fast running when a number of successive legs are off the ground (Part 8, pp. 299, 304). This requirement is absent in Diplura. The dorsal system of the japygids is much more complex than that in the campodeids because of the presence of intercalary tergites, and the japygid musculature provides strength and flexibility of trunk. The proturan dorsal system is more complex than in Cizmpodea and fans out posteriorly (Prell, 1913, P1. 11, Fig. 8). (ii) Ventral longitudinal muscles In Campodea the very simple series of ventral muscles linking the tendons above the presternite bulges and the endoskeletal plates has been noted above. Anteriorly the prothoracic ventral longitudinal muscles insert upon the median head endoskeletal tendon system (shown in blue, Manton, 1964, Fig. 47). In Japyx the ventral longitudinal muscles are complex, providing both strength and flexibility between the thoracic segments. The spine apodeme interrupts linear continuity and provides great length to certain sectors (e.g. vzm.2., figure 10). A strong series of muscles pull on the posterior margin of each thoracic sternite, vlm.3., vlm.4., vlm.5. (figure 10). Only vzm.1. forms a
79 EVOLUTION OF HEXAPOD LOCOMOTORY MECH+NISMS 281 continuous band linking successive lateral endoskeletal tendons, it is the most lateral sector and continues into the abdomen, missing out attachment at the metathoracic-first abdominal intersegment. Sectors vlm. 3. and vlrn. 4. insert respectively on the sternal cuticle between the arms of the Y-skeleton and lateral to them (see mesothorax in figures 10, 11F, G). Sectors vlm.5. and vlm. 6. insert on the lateral endoskeletal tendons. Sector vlm. 7., from the lateral endoskeletal tendon, passes backwards to the lateral furrow between the presternites and is indicated only in part in figure 10, prothorax. Thus this ventral longitudinal muscular system of japygids is particularly suited to provide strength and thoracic flexibility. In the Protura the ventral muscles are simple (Berlese, 1910 Pls. X, XI, text-fig. V) but with some long sectors in the thorax. (iii) Deep dorso-ventral and deep oblique muscles This series of muscles provides stability in Chilopoda and Pauropoda, while the superficial pleural muscles are largely concerned with trunk flexibility. The latter are absent in Pauropoda, where rigidity is the principle need, and are present in Symphyla, where flexibility is the main requirement, the deep dorso-ventral and oblique muscles here being absent (Parts 8, p. 273 ; 9, p. 138). The Diplura show marked thoracic flexibility but also need stability for the long (Campodea) or strong (Japyx) legs, and both the deep dorso-ventral and deep oblique muscles, and the superficial pleural muscles are well developed. The deep oblique muscle dvmd. arises in Campodea from tendons anchored between the presternite bulges on the meso-and metathorax and from the tentorium*. The three muscles slope backwards and upwards to the tergites of the succeeding segments, as shown (figure 13a), thereby gaining great length (cf. Chilopoda, Part 8, figures 50, 51 where these muscles do not leave their segments and figures 60, 67, 78 where they insert on the following segments). In Japyx these muscles arise from the postero-ventral cranial margin and from the posterior margins of the pro- and mesothoracic sternites near the middle line, dvmd. (figure 10). The muscles pass up and back to insert on the anterior margins of the following intercalary tergites ending on the tendinous junction between principal and intercalary tergites, as shown. Stout deep dorso-ventral muscles dv. pass from an endoskeletal unit upwards to the following tergite in both Campodea and Japyx (figures 12F, 13a, lo), pulling directly or indirectly on the apodemal skeleton. A single sector leaves the edge of the median endoskeletal plate in Campodea and a fan of subsidiary sectors, dv.l., dv.2., dv.3., with dv. leave the lateral endoskeletal tendon in Japyx, inserting on the tergite above, on the pleuron and on the following intercalary pleuron (see meso- and metathorax in figure 10, 11F). A third muscle, dvc. is present in Japyx only. At the articulation of the coxa a tendon passes upwards from the arm of the Y-skeleton and forms the origin of muscle dvc. (see pro- and mesothorax on figures 10, 11E). This muscle is strikingly similar to that of the Scolopendromorpha, and probably serves similar needs (Part 8, p. 275) namely stability of leg base in presence of much trunk flexibility, see also muscle tl of Symphyla (Part 9, p. 119, figure 5). In Manton, 1964, figure 47 this prothoracic dvmd muscle is unfortunately labelled lower sector of dorsal longitudinal muscle.
80 282 S. M. MANTON In the Protura there are no typical antagonistic deep dorso-ventral and deep oblique muscles (see muscles dv and ob in figure 12a) which normally promote rigidity in other classes. Berlese s intersegmental dorso-ventrals (see his text-fig. IV) probably promote flexibility, such as is conferred by the superficial pleurals of Chilopoda and Diplura and the endoskeletal muscles of Diplura. Muscle ilo of Berlese from the posteremedian part of the sternite to the antero-lateral part of the tergite two segments behind is a little like the japygid muscle dvmd. (figure 10) and muscle dpll. of some Chilopoda (Part 8, figure 51). (iv) Endoskeletal muscles In the Diplura these muscles appear to be mainly concerned with general stability and with facilitating intersegmental flexures, constricting the intersegmental regions. This category of muscles is absent from myriapods because suitable endoskeleton is not present (except perhaps for muscle sp. of Symphyla (Part 9, figure 5)). In Gzmpodea an oblique endoskeletal muscle 1 passes from the lateral endoskeletal bar to the trochantin and furrow between it and pleurite 2 (figure 12E). Oblique endoskeletal muscle 2 from the middle of the endoskeletal plate inserts on the margin of the following tergite (figure 12F). Oblique endoskeletal muscle 3 is more posterior, leaving the side of the endoskeletal plate, along with a stouter transverse endoskeletal muscle to the pleuron (figure 12c). In Japyx a similar transverse endoskeletal muscle is present (figure 1 lg, and foreshortened on mesothorax in 10). There are no muscles exactly like the three endoskeletal muscles of Gzmpodea, but possibly their homologues are dv.i., dv.2., dv.3. in Japyx, whose tensions must have roughly similar effects. The evolution of these endoskeletal muscles in the Diplura is dependent upon the presence of the endoskeleton, and this has been necessitated by the formation of either long or very strong legs in these hexapods in contrast to the myriapods. In the Protura Berlese (1910 text-figs. IV, P1. XI, fig. 19) figures a transverse muscle ts spanning the anterior part of the meso- and metathorax which is presumably similar in function to the transverse endoskeletal muscles of Diplura (figures 10, 12). (v) Superficial pleural muscles The presence of these muscles in Diplura is associated with marked thoracic flexibility, as in the trunk of epimorphic Chilopoda and Symphyla (Parts 8, p. 273; 9, p. 119). In Cizmpodea the superficial dorso-ventral muscles 1 and 2 (figures 12G, 13b) arise respectively from the antero-lateral corner of the sternite and from the antero-lateral margin a little behind. The former passes upwards and forwards to insert on the cranium or on to the anterior parts of the meso- and metathoracic tergites, and the latter pass upwards and backwards to the
81 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 283 overlying tergites. A pleural muscle (so marked on figure 12E) in Curnpodea arises as two sectors, one from the trochantin and one from its junction with pleurite 2, and passes upwards and inwards to the middle of the anterior edge of the succeeding tergite. This muscle and oblique endoskeletal muscle 2 must assist coxal levation by lev. co. In Jupyx there are three pairs of superficial pleural muscles (figure lo). A superficial dorso-ventral a, from the postero-lateral corner of the meso- and metathoracic sternites, inserts on the tergite of the succeeding segments. A substantial superficial dorso-ventral b, from the anterior border of the procoxa and lateral parts of the presternites, inserts on the principal tergite, just behind its intercalary junction. A very large superficial pleural muscle (so marked on figure 10, meso- and metathorax), forms a fan from the intersegmental pleuron to the preceding tergite. On the prothorax there is a slender anterior dorso-ventral muscle, drawn but not labelled on figure 10, and from the small Y-shaped apodeme anterior to that bearing the spina and coxae, a stout muscle passes upwards to the cranium. The ventral origin of this muscle is cut in figure 10. The Protura show no such category of muscles, their trunk flexibility being catered for by Berlese s intersegmental dorso-ventrals, see above. 8C. CONCLUSIONS CONCERNING THE THORACIC MUSCULATURE OF THE DIPLURA AND PROTURA The conspicuous flexibility of the thorax of the Diplura is differently contrived from that of the trunk of epimorphic Chilopoda and Symphyla, although all three classes possess a flexible pleuron and intercalary tergites. The intersegmental flexibility of the dipluran thorax is due to segment length, to lateral constriction by transverse endoskeletal muscles and to strong flexures caused by the other endoskeletal and pleural muscles. Corresponding movements in Chilopoda and Symphyla depend upon long and oblique muscles dpl. and dp2z. in a lateral position pulling from the segmental tendons (Part 8, figures 50, 5 1, 60). Chilopodan and symphylan superficial pleural muscles differ in detadfrom those of the Diplura, the resemblances being only superficial. The deep dorso-ventral and deep oblique musculature of Diplura also differs from those of myriapods although these categories of muscles confer stability in those classes in which they are present. The dipluran muscle dvmd. is roughly like muscle dvma. of Chilopoda and Pauropoda (Parts 8, figures 50, 51,60; 9, figure 8a) and there are no dipluran muscles corresponding with dvmp. and dvtr. of Chilopoda and Pauropoda. Muscle dv. of the Diplura is not like any myriapodan muscle. Thus the Diplura differ from the myriapods in the details of their musculature, the partial similarity in the general lay out being correlated with similar needs. The small size of the Protura would be expected to lead to simpler and fewer muscles than present in the larger Diplura, as indeed is the case. But there is much difference in detail, and the general similarities existing between the proturan, dipluran and myriapodan musculature are convergent and associated with needs. A similar conclusion was reached concerning the general form of the exoskeleton, pleuron and mode of action of the leg-base.

82 2 84 S. M. MANTON coxo-rternne articulation a axis of swing of cox2 -. EOSE NTOMON C 3 b

83 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 285 Figure 9. Heterojapyx novaezeelandiae Verhoeff. External side view of part of the thorax to show the trunk and limb sclerites, the limb base and the axis of swing of the leg on the body, the head end is to the left. The most sclerotized cuticle is mottled and the more flexible pleuron and arthrodial membranes are white. The edges of most sclerites are not sharply defined but merge or fade into the more flexible surrounding cuticle. Leg 1 is turned forwards and leg 2 backwards at the coxa-body joint, the axis of swing being shown by the heavy vertical line. The articulations, or corresponding points of close union, between sclerites are marked in black. Heavy sclerotization strengthens these points: the ventral coxa-sternite articulation; the anterior and posterior components of the coxa-trochanter pivot articulation; the anterior component of the trochanter-femur pivot articulation, the posterior component being absent; the femur-tibia pivot articulation; and the dorsal tibia-tarsus hinge. The coxa-trochantin union consists of a coxal lobe, not heavily sclerotized, which is tucked under the edge of the trochantin and linked with it by ample arthrodial membrane, the morphology here strengthens the coxal movement and limits it to a promotor-remotor swing about line shown. The labelling of the sclerites is intended to indicate the similarities and differences between those of the Chilopoda (Part 8, figures 1, 41, 45a, 48, 49, 58, 65, 73, 74, 75). Entomological custom terms the intercalary tergite of the meso- and meta-thorax and a precosta and that of the prothorax a pretergite. The chilopodan procoxa corresponds with the entomologists merosternite. Pleurite 2, the katopleure of entomologists, does not correspond with the chilopodan katopleure. (b-c) The proturan Eosentomon, after hell (1913). (b) Lateral view of the mesothoraic segment, the coxa being seen end on. Head end to the right. The positions of the two coxal articulations are marked by black spots. The axis of the promotor-remotor swing of the coxa is marked, and the coxal rock, caused by a sliding of the trochantin (pleurite 1) against pleurite 2 at the sliding facets drawn by Prell (pleural articulation), is shown by the arrows within the coxal cavity. The three arrows, dorsal and ventral, mark the inter- and intrasegmental levels of flexibility. (c) Ventral view of mesothoracic segment to show the subdivision of the sternite, into the zones acro-(ac.), pre-(pr.), meso-(ms.) and meta-(met.) sternite of Prell, the position of the coxasternite articulation and the median sternal apodeme. Parts of pleurites 1 and 2 (pls. 1, 2) are visible. The horizontal arrows mark the levels of inter- and intrasegmental flexibility.

84 JAPYX cranium Dr.co ,... I - intercalary tergite 3 transverse endoskeletal muscle transverse endoskeletal muscle 1 1 superficial dorm-ventral b. intercalary tergite \ superficial dorso-ventral b superficial pleural muscle I woerficlal DiPIIT.1 mll<cie abd. I tergite abd I arm of Y ridge / c r mm. Figure 10. Japygid sp. View, from the sagittal plane of the principal muscles of the three thoracic segments, numbered above. Fat body and vascular system are omitted and endoskeletal tendons are shown in black. The levels of the transverse sections shown in figure 11 are marked by E, F, and G below. The ventral Y-shaped ridge (see figure 9) consists of a median infolding of cuticle from which arises the median spine-like apodeme, the paired arms appear in internal view, stippled white. The prothoracic segment is cut back as far as the base of the coxa, muscle lev.co. and the lateral endoskeletal tendon being cut short. These structures are intact on the mesothoracic segment. On the metathoracic sep'nt the median spine-like apodeme and its appended muscles are cut away in order to expose the ventral longitudinal muscles. The metathoracic ganglion is shown, but the nerve cord is removed elsewhere. Three muscles are inadequately shown: muscle vlm. 7 arises laterally, mainly from the furrow between the presternites and from the procoxa, and passes forwards to the lateral endoskeletal tendon of the preceding segment (see prothoracic segment); another muscle from the same origin passes obliquely upwards and forwards to the anterior margin of its own tergite at the tergite-pretergite junction, not shown; and the superficial dorsci-ventral muscle b arises largely from the procoxa, but also from the junction of the tergite and posterior presternite and from the posterior presternite itself. The transverse endoskeletal muscle, shown much foreshortened in the metathoracic segment, arises, as shown in figure 11G, from a median sheet of connective tissue extending upwards from the spine apodeme which cannot be drawn in sagittal section without obscuring muscles vlm.2 etc.

85 E, dorsal longitudinal muscles F Intercalary tergite 3 G ntercalary rergtri 3 transverse _endorkeleul muscle cuticle coxo- sternal articulation \ / lacera: endoskeletal tendon lateral arm of Y ridge vlm.4. median arm of Y ridge COX2 1 Figure 11. Japygid sp. Thick transverse sections, viewed from in front, at the levels E. F, G marked on figure 10, to show the principal features of the trunk musculature and extrinsic muscles of the leg. The anterior margin of the mesothoracic coxa is cut in E, the posterior part of the coxa is cut in F and in G the coxa is free from the body. Section E cuts through the tip of the spine apodeme from the prothorax, marked cuticle and surrounded by ventral longitudinal muscles 2 above, and laterally and ventrally by the enormous coxal retractor muscle 6. Section F passes through the ventro-lateral tendon extending inwards from the arm of the Yskeleton of the mesothorax, here an infolded ridge marked lateral arm of Y-ridge. Section G passes through the origin of the spine apodeme of the mesothorax. showing a median tendon above it, on to which are attached the pair of transverse endoskeletal muscles foreshortened in figure 10. I

86 288 S. M. MANTON
87 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 289 Figure 12. Curnpodea sp. Diagrammatic thick transverse sections to show the skeletomusculature of thorax and legs, the fat body is omitted. Cut muscles are rulled, nerve cord and ganglia are stippled, and cuticular endoskeleton is white. The levels E, F, G, EE, FF of the sections are marked on figure 13. (a) Anterior view of the prothorax and appended leg. On the left (E) the anterior face of the trochantin (mottled) is shown overlying the coxa, but the sectional outlines of both coxa and trochantin are drawn as if visible through the trochantin. The protractor muscles to the coxa are shown on the left. On the right (F) the same outline of the coxa is shown but the retractor muscles are drawn. The section of the body is a little more posterior than in (E), no pleurites are cut and, looking into the cavity of the piece of trunk, the anterior edge of the tergite of the second thoracic segment is visible. (b) Anterior view of the body at level G behind F (see figure 13a) showing the anterior part of the mesothoracic tergite and the median endoskeletal plate behind the prothoracic legs. The slope of superficial dorso-ventral muscles 1 and 2, the former arising from the anterior edge of the mesothoracic sternite and the latter arising more ventrally from this sternite, are shown in figure 13b. (c-d) Anterior views of the ventral parts of the mesothoracic segment corresponding with those of the prothorax in (a), to show: the differences in the course of coxal protractor muscle, pr.co.3.. which inserts on the mesothoracic tergite above instead of upon the median endoskeletal plate as in the prothorax, see (E); the additional coxal protractor muscle, ~r.co.4.~ arising from the anterior margin of the coxa; and the differences in the endoskeletal connectives, the skeletal struts from sternite 2 to the lateral endoskeletal bars arising laterally instead of mid-ventrally as on sternite 1, cf. (E), in (d) there is a stout fork from the median plate to the sternite which is absent on the prothorax, and behind this fork the median endoskeletal plate is united with the sternal cuticle as shown in (b) for the prothorax. These three types of connection with the cuticle are also shown in figure 13a.
88 290 S. M. MANTON
89 Figure 13. Cumpodeu sp. Longitudinal views of the thoracic musculature, conventions as on figure 12. (a) Internal aspect of the right half of the animal showing the dorsal and ventral longitudinal muscles, the deep dorso-ventral (dv.) and the deep oblique (dvmd.) muscles and the endoskeleton. The sternites are well separated by arthrodial membrane and the ventro-laterally situated bulges of the body wall, at the anterior ends of the meso- and metathoracic segments, corresponding with presternites 1 and 2 of Jupyx (figure 9). are shown by dotted white lines. From the furrow between these bulges an intersegmental tendon runs into the ventral longitudinal musculature and forms the site of origin of muscles dvrnd., passing to the tergite of the segment behind. The very long muscle dvrnd., inserting on the mesotergite, arises from the tentorial apodeme (see Manton, 1964, figure 47, where this muscle is unfortunately labelled dorsal longitudinal muscle (lower sector) ). The ventral longitudinal muscle is more complex than drawn and is linked with the lateral endoskeletal bars, the median endoskeletal plate, the intersegmental tendons between pro- and mesothorax and between meso- and metathorax (mentioned above), and anteriorly the ventral longitudinal muscle inserts on a median endoskeletal bar (shown on the left) which is linked by fibres to the neck intersegment as shown, and anteriorly ends on the endoskeletal plate in the head. The posterior median end of the endoskeletal bar in the head is shown in blue in Manton, 1964, figure 47, the more anterior part being labelled paired endoskeletal connective. The median endoskeletal plates of all thoracic segments bear antero-lateral extensions, the lateral endoskeletal bars (seen more clearly in figure 12e); the latter are united with the sternite below by paired struts, labelled on the prothorax. A stouter median Yshaped strut follows on the meso- and metathorax (see figure 12d), and behind this level on all thoracic segments a long median infolding of cuticle, marked by white striations, and better seen on figure 12b, supports the median plate. There are no more posterior deep dorso-ventral or oblique muscles. (b) Internal view of the right half of the thorax, the muscles and body wall shown in (a) being cut away to expose the extrinsic limb muscles and the superficial dorso-ventral muscles. The coxae are cut to show the origins of the protractor muscles pr.co.z., ~r.co.2.~ and pr.co.3., and the retractor muscles ret.co.i., ret.co.z., rer.co.3., and ret.co.#., which, as shown, are not exactly the same either in number or disposition on the three segments. Muscle ret.co.4, inserts on to the anterior margin of the prothoracic sternite.
90 292 S. M. MANTON posterior d Figure 14. The femur-tibia joint of the Diplura. (a-b) Distal views of the end of the femur of Heterojapyx nouaezeelundiae Verhoeff, (a) with the tibia downwardly flexed on the femur, and (b) with the tibia much more depressed, as indicated by the solid arrow on (c), so exposing the dorsal process 2, bearing the tendon of the tibial extensor muscle, lev.ti., see (c); the points X and Y marked on both (a) and (b) show the movement. A very strong femoral anterior component of the pivot joint is marked W, there is no posterior component. The right-hand arrow on (b) shows the direction of rotation of the femur caused by contraction of the femaral rotator muscles ro+fe.,, (see figure 12F); this rotation is transmitted to the tibia (lower arrows) by the articulation between processes Wand 2 at the femur-tibia joint. (c) Posterior view of the femur-tibia joint of Campodea, its structure being essen&ially as in Heterojapyx (a, b). The femoral part of the anterior component of the pivot joint lies on the anterior side of the tibial process Z and is marked by the dotted line W. There is considerable emargination of the distal rim of the femur on the posterior side. Only the antagonistic pair of muscles operating the femur-tibia joint (lev.?i and dep. ti. ) are shown, other muscles traversing this joint (see figure 12F) are omitted. (d) Posterior view of the tibia-tarsus hinge joint of Campoden
91 EVOLUTION OF HEXAF'OD LOCOMOTORY MECHANISMS TRUNK MORPHOLOGY AND JUMPING MECHANISMS OF THE APTERYGOTE CLASSES The employment of jumping gaits, or large jumps as escape reactions, has been independently acquired by different means in the Collembola and Thysanura. The jumping mechanism of Collembola appears to be a fundamental attribute of the class, which has controlled the evolution of the trunk morphology. Jumping has secondarily been lost in many Collembola with consequent modification of the basic structure. The Machilidae also utilize a well developed, but quite different abdominal jumping mechanism for escape reactions. The collembolan trunk morphology, internal and external, has been recorded below as far as needed to explain the jumping mechanism. The cuticle, endoskeletal tendon system and the trunk musculature of Tomocerus are described here because no existing accounts of any collembolan are sufficiently accurate or detailed as to provide a basis for an understanding of the functional significance of even the gross externally visible morphology of these animals. Endoskeletal systems of several apterygotes have been described in some detail by Barlet (1951, 1967), Carpentier (1949), Barlet & Carpentier (1962), but without reference to the animals' movements. Muscles of Collembola have been described by the above, inaccurately and in part by Maki (1938) and Lubbock (1873), and false interpretations have been placed upon these findings by writers of textbooks and monographs. The wide pleuron and laterally projecting coxae seen in the Diplura and Protura are absent in Collembola and Thysanura, where the coxae are ventrally projecting from the body and the tergites embrace the whole trunk as far as the leg base. The jumping Collembola and machilid Thysanura have particular need for boxing in the body in their control and use of changing internal hydrostatic pressure during jumping. The form of the thoracic endoskeleton and musculature is affected by the tergite envelopment. 9A. THE COLLEMBOLAN JUMP The jumping mechanisms used by the terminal springing organ is one which has affected the entire construction of the body and legs. The entomological monographs and textbooks state that the propulsive thrust of the springing organ is caused by extrinsic muscles appertaining to that organ, and that antagonistic muscles account for the rest of the movements. In fact there are no extrinsic muscles from the springing organ which are directly propulsive. All such muscles are concerned with recovery movements and with stabilization. The propulsive thrust of the springing organ against the substratum is almost entirely hydrostatic in origin, aided by cuticular elasticity and some intrinsic muscles acting on the distal furca or dentes of the springing organ. The volume of the trunk haemocoel is large compared with that of the springing organ. A small longitudinal contraction of the body, effected by very stout and strongly constructed musculature, gives a sudden hydrostatic force blowing out the springing organ and swinging it strongly away from the body. The cuticle and joints of the whole body are constructed so as to resist any tendency towards dilatation elsewhere under such pressures. Typical arthrodial membranes, such as usually present in arthropods, are absent. It has been shown by Noble-Nesbitt (1963) that desiccation stops the springing movements of Podura, a sufficiency of haemolymph pressure being essential for the
92 294 S. M. MANTON working of the spring and for eversion of the ventral tube. These findings are in keeping with the present account of the spring mechanism. (i) The cuticular connections and movements of the springing organ of Tomocerus The position of the springing organ of Tomocerus at the end of a jump, and the resting position when the manubrium is flexed forwards, are shown in figures 15a and b respectively. The manubrium of the springing organ is closely articulated posteriorly with a thick transverse sclerotized bar situated at the ventral rim of tergo-pleural arch 5, shown by a spot on figures 15a, b, and as a double layer of thick cuticle in the transverse section in figure 27U. These two cuticular rims are united by a short link of thinner cuticle, and together they form a very strong transverse hinge about which the manubrium swings in the median plane. But the whole joint between manubrium and body is as remarkably constructed, and serves as special a purpose, as does the coxa-trochanter joint of a chilopod leg (Part 8). The ample expanse of arthrodial membrane, which is normally present on the opposite side to the actual hinge of a hinge joint (Part 6, figure 14 and Part 8, figure 63 femur-tibia joint, etc.), is absent between the manubrium and the body. Instead there is an astonishing array of sclerites and mobile junctions. Antero-ventrally the manubrium articulates on either side of the mid-line with a pair of strong conyles formed by the ends of paired, thick, highly sclerotized basal rods which traverse the median basal plate 4 below tergo-pleural arch 4 (figure 15). The articulation of one side is marked by a cross on figures 15a-c and both articulations are marked on pl. 4, figure c. Each consists of wide facets of highly sclerotized cuticle separated by a short link of thinner cuticle. The median basal plate 4 on figure 15a-c embraces most of the sclerite termed ventral plate by Dennis (1949). It lies below tergo-pleural arch 4 from which it is separated by a furrow (figure 16 ab4, e) of thinner cuticle, shown as a line on pl. 4, figure d between BP4 and 4, and extending forwards as far as the hamula. Basal plate 4 is infolded ventrally to form a median gulley into which the manubrium can be flexed at rest (figure 16 ab4 and pl. 4, figures a, c, g). The ventro-lateral angle of the basal plate forms a conspicuous posterior lobe marked p on pl. 4, figures a, c, d. Postero-laterally basal plate 4 is separated from paired basal plates 5 by shallow vertical intuckings of flexible cuticle on either side (figure 15a), and the letter f lies over this junction in P1. 4, figure c. Basal plate 5 is triangular in shape, its ventral point passing uninterruptedly into the sclerotization of the condyle on the basal rod on either side. Posteriorly basal plate 5 narrows to end at the posterior hinge of the manubrium, and its margin with tergo-pleural arch 5 is a simple hinge line (figure 160) which projects slightly outwards and not inwards. The proximal part of the manubrium, marked basal sclerite on the figures, is wide (figure 27T-U). It forms a flexible junction with the ventral border of basal plate 5 on either side and carries some proximal slots of lesser sclerotization, marked by the two right hand f on figure 27U, the ringed point n is shown in section in figure 16n. The middle of the manubrial rim at the posterior hinge is also interrupted by a similar break in the high degree of
, where this feature assists in the bending of the cuticle.")
93 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 295 sclerotization, (left hand f on figure 27U). The slotting in the cuticle is directly comparable with similar, but less elaborate, slotting in the trochanter of a scorpion (Manton, 1958a), where this feature assists in the bending of the cuticle. When the manubrium is extended, all the parts just described can be seen. The resting position of the basal plate 4 is as shown in figure 15a, where the basal rods are approximately straight, and their cuticle is under no outside tension. But when the manubrium is flexed forwards under the abdomen about the posterior hinge, the articulations are pressed deep into the body, as indicated in figure 15b. The basal rods are then greatly curved, and basal plates 5, together with the slotting in the base of the manubrium, allow sufficient distortions for this remarkable intucking to take place. The outwardly directed hinge line between basal plate 5 and the tergo-pleural arch above (figure 15) and the inwardly directed junctions between basal plates 5 and 4, assist in the infolding of the ventral parts of basal plates 5. The movement is effected by muscles (see below), and when completed the hamula impinges into the well-known slot at the base of the dentes (furca), figure 17V, unlabelled, and the muscles can then relax. But the bending upwards of the posterior ends of the basal rods leaves their cuticle in a state of tension, which, when released by the disengagement of the hamula and dentes, assists the backward and downward movement of the manubrium at a jump. A very wide angle of movement (some 120 ) of the manubrium on the body is made possible by these cuticular arrangements and they are such as to withstand sudden and great changes in internal hydrostatic pressure. Ample arthrodial membrane at this joint might provide a fairly wide angle of swing, as at the coxa-body joint of Scolopendra or Lithobius, but would be quite unsuitable for a hydrostatic mechanism because ballooning outward of such an arthrodial membrane would weaken the hydrostatic force transmitted to the manubrium. There are other examples of hinge joints which are elaborated to give wide angles of swing, e.g. the femur-patella joint of a spider (Manton, 1958a), and the thoracic legs on the body of a copepod (Perryman, 1961) the coupler in the latter providing strength and the great mobility. The most free of myriapodan hinge joints (Parts 6, 8), with considerable emarginations of the podomeres and am le arthrodial membranes, do not permit anything like the wide angles of P 100 found in the movement in the spider leg or the 120 by the collembolan manubrium. Typical arthrodial membranes are unsuitable not only at the manubriumbody joint, but everywhere in the body, because they would tend to balloon outwards on increase of hydrostatic pressure. The absence of such membranes (figure 18a, c/f, Part 5, text-fig. 80, am, Parts 7, figures 3, 6, 7, 10, 14, 17-22; 8, figures llc, 13, 15, 76) is just as much an integral feature of the jumping mechanism as are the features of the manubrium-body joint. There is conspicuous lack of intersegmental flexibility of trunk in the jumping Collembola in contrast to the more flexible hypogastrurids, Neunuru (pl. 3, figures h, i). In the former each tergo-pleural arch is connected to the next by a short link of thinner but well sclerotized cuticle (figure 18a), while the latter show much more flexible cuticle in this position. Elasticity of the tergo-pleural arches themselves probably aids the recovery movement after a jump. These arches are bare of muscle insertions on their
94 296 S. M. MANTON faces, apart from the deep dorso-ventral dv and the superficial pleural muscles (figures 20a, 21a), in contrast to the many muscles inserting on the faces of the tergites of Chilpoda, etc. (Parts 8, figures 55, 61, 62; 9, figures 4, 8). Slight shortening of the body caused by contraction of the longitudinal and deep oblique muscles (figures 19, 20b, 21) probably arches (corrugates) the tergo-pleural sclerites slightly. Extension of these relaxed muscles at the end of a jump will be mediated largely by the deep dorso-ventrals inserted upon the face of the tergal regions and partly by elasticity restoring the sclerites to their resting shape. (ii) The extrinsic muscles, and their functions, of the springing organ and hamula of Tomocerus The extrinsic muscles of the manubrium form two groups, those concerned with strongly flexing the manubrium under the abdomen after a jump, and those mainly concerned with the maintenance of stability of the manubrium. The muscles are shown diagrammatically in figure 17a. The largest muscle, marked 1, arises from the proximal antero-lateral part of the manubrium. The muscle passes forwards through abdominal segments 4 and 3 to end superficially on connective tissue at the abdominal intersegment of tergo-pleural arches 2 and 3. The origin of the muscle is shown in figure 27T, where the position is ringed, and the detail is shown at a higher magnification in figure 17c. Here an infolding of thin, but considerably sclerotized, cuticle carries a wide area of tonofibrils supporting the muscle. This muscle junction is flexible because the intucked cuticle is thin; the junction cannot be everted or stretched under internal hydrostatic pressure; but it can open out a little when muscle 1 is relaxed, so aiding in small measure the wide angle of swing of the manubrium. But the principal service of such a junction is probably a swinging of the intucking about the manubrial hard parts when the manubrium is in different positions, so enabling muscle 1 to pull on its tonofibrils in a constant direction, without change of angle between the two. This flexible intucking serves exactly the same purpose as the arcuate sclerite in the femur-patella joint of the spider leg (Manton, 1958a, p. 172), namely the maintenance of a constant direction of pull of the muscles on their tonofibrils throughout a wide angle of swing of the distal parts, the patella in the spider and manubrium in Collembola. The course of muscle 1 is seen in sections N to S figures 26, 27, and the insertion in section M, figure 26. Contraction of muscle 1, when the manubrium is outstretched, cannot do anything other than strongly flex the manubrium forward about the posterior hinge, marked by a spot on figure 15a but unmarked on figure 17a. This muscle cannot extend the manubrium. Each postero-lateral corner of the posterior hinge between manubrium and body bears an inwardly directed tendon 4 marked on figures 27T, 21a but unlabelled on figure 17a. This paired tendon bears several muscles. Muscle 6 is substantial, and inserts on either side on the upper part of the intersegment between abdominal tergo-pleural arches 3 and 4. The origin and insertion of the muscle is shown in figure 17a and it is cut transversely in figure 27s. A fan of smaller muscles, s, t, u also originates on tendon q and passes backwards to the anterior margin of the 6th abdominal segment, as shown in figures 17a, 21, 27T-U where muscle t is seen to be the bulkiest of the three, its origin showing in T and the muscle is cut in U.

95 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 297 A fine paired tendinous strand dv (figure 21a) leaves the tendon q and passes upwards to end on the tergo-pleural arch above by tonofibrils. It bears no muscles. Two more extrinsic manubrial muscles are closely associated functionally with the muscles arising from tendon 4. They are the substantial muscles ex, 1. and ex.2. arising from the manubrium close to the hinge, as shown in sections T-U Figure 27. They pass upwards and forwards to insert on the antero-lateral face of tergo-pleural arch 4, see figures 17a, 27s. Since extrinsic muscles ex. 1. and ex.2. arise close to the hinge, they cannot cause either flexure or extension of the manubrium. These muscles, together with muscles 6,s,t,u and tendon dv, described above, can all contribute to maintaining firm alignment of the manubrium on the body during the jump. They will also counter any tendency for the manubrial hinge to be blown outwards from the body when internal hydrostatic pressure is greatest and forcing the manubrium to swing downwards and backwards. These muscles are probably mainly stabilizers, but their size and strength is such that they may pull the whole hinge inwards into the body slightly during a jump, deforming the body sclerites in the vicinity. Such a movement would contribute to the width of the angle of swing of the manubrium. The homologies of the extrinsic muscles of the manubrium are plain. Muscle 1 is an elaborated homologue of component 1 of the typical lateral longitudinal muscle system of other segments (figure 20b). Muscle 6 represents some of the other components of this system, which are not serially recognisable in segment 4. Tendon dv appears to correspond with deep dorso-ventral muscle dv of other segments and to represent this muscle on segment 5 (figure 21a). Muscles s,t, and u correspond with the deep oblique muscles ob of all trunk segments, except abdominal 4, see figure 21a. Muscles ex.l. and ex.2., arising within the proximal edge of the manubrium, appear to represent the extrinsic limb muscles of a limb-bearing segment, and do not represent any part of the trunk muscular system. The hamula presumably undergoes minute movements resulting in its fixation into the notch at the base of the dentes (figures 17a, 27P, Q) when the springing organ is folded forwards and at rest, and release from the notch at the jump. It is probable that the hamula can be inflated slightly and extended by haemocoelic fluid when its extrinsic muscles relax. The ventral tube is extended in this manner. The many muscles of the hamula are described in Appendix I (vi) and shown in figures 27P, Q, 17a, 21a. The elaborate arrangement of small muscles, with tendinous junctions, cannot do other than cause small movements in various directions, and muscle F must be able to shorten the hamula and pull it forwards slightly and out of the notch, tension on the other muscles being appropriate. (iii) The intrinsic muscles, their tendons and functions, of the springing organ of Tomocerus The dentes or furca is attached to the manubrium by a pivot joint with a small range of movement, a strong articulation between the two being situated laterally. Two pairs of tendons traverse the length of the manubrium (figure 17a). Tendon t carries no muscles. Distally it spreads out over the firm
96 298 S. M. MANTON internal surface of the epithelium lining each tubular dens, and in particular forms a strong connective tissue support round the small pore leading from the internal cavity of the manubrium to each dens. One pore is shown in longitudinal section in figure 17b. Tendon t lies laterally in the manubrium, as shown in figure 17W, and ends proximally on the cuticle at the margin of the manubrium (figures 17a, 27T). This tendon must prevent the dentes from being unduly inflated by internal pressures from the manubrium, and any tendency towards dislocation at the manubrium-dentes articulations is thereby controlled. The highly sclerotized and thick manubrial cuticle is differentiated on the anterior and posterior faces. Postero-dorsally there is a deep longitudinal groove of thick highly sclerotized, and therefore elastic, cuticle (see figure 17W, the square p being shown in section at a higher magnification in figure 16p). The shoulders of the furrow are formed of thick cuticle staining pink with Mallory, as opposed to the red-staining lateral cuticle shown at Y figures 17W, 16r. This pink cuticle is also seen at the bases of the dentes, figure 17V and is probably more elastic than the red. * On the anterior face of the manubrium is a highly sclerotized longitudinal hinge in the form of a shallow groove containing a short link of very thin, highly sclerotized cuticle which will allow slight opening out and the reverse (figure 17W). Two pairs of muscle sheets lie within the manubrium. Muscle sheets ma arise from a long tendon u which pulls on the posterior margin of each dens causing a backward swing of the dens about its pivot joint with the manubrium. Muscles ma pass obliquely in a proximal direction from the tendon to insert all along the sides of the postero-dorsal groove as shown. These many, but short, muscle strands are suited to exert a large tension on the tendon but to cause only a small displacement. Such an arrangement must cause pressure by the dentres on the substratum in addition to the force set up by the swing of the manubrium. These muscles mu contrast with the flexors to the claw tendon of so many arthropods. Where the tendon is long the claw flexor muscles are also long, and the resultant movement of the claw is considerable, strength being obtained by the presence of many long sectors of this muscle system inserting on many or all of the leg segments (Part 6, figure 6 Pun. t. and associated muscles, also figures 4, 17, 19-21; Part 8, figures 53, 63). Only in Scutigera are some of these muscles short, forming a series within the tibia resembling muscle ma in Tomocerus (cf. Part 8, figures 72a, c). This feature in Scutigeru is associated with the presence of a multi-segmented tarsus, many joints contributing to terminal flexure of the leg, and the need for great strength by the claw. But long muscles are also present here extending to the prefemur. Paired muscles mb. span the internal cavity of the manubriurn of Tomocerus. They are widely attached antero-laterally and slope obliquely in a distal direction, opposite to that of muscles mu, to insert in the middle of the postero-dorsal groove (figure 17a and W). Contraction of these muscles must deepen the dorsal groove, so bending the entire manubrial cuticle, which will regain its shape by elasticity as soon as muscle tension is released. The volume of the manubrium will be slightly reduced by contraction of musclesmb. The * The mechanical properties of cuticle differentiated by Mallory s triple stain were considered in Part 5, p. 174.

97 EVOLUTION OF HEXAPOD MCOMOTORY MECHANISMS 299 function of muscle mb must be twofold. To brace the manubrium against the increasing hydrostatic pressure from the body during the jump and to increase this hydrostatic thrust on the dentes by deforming the manubrium, so pressing the dentes hard against the substratum. A springing organ in the form of an undivided rod would not jump quite so strongly as does the two-segmented organ, and the double ended dentes gives a slightly flexible structure which makes for stability in the take off on rough substrata, besides increase of thrust against the ground. (iv) The trunk musculature of Tomocerus The sudden, slight, shortening of the body, which creates the momentary pressure needed to work the springing organ, is mediated by cuticular and skeleto-muscular features. The musculature of both thorax and abdomen creates the hydrostatic pressure. The hamula and its connections holds the springing organ in place until muscular tension is sufficiently built up, and then the whole apparatus is released, resulting in the jump. A summary of the trunk muscles of Tomocerus is shown by figures 19, 21 and transverse sections at the marked levels A-U are given in figures Muscle details and functions are described in Appendix I. A functional analysis of the musculature of the several classes of Arthropoda so far considered (Parts 6, 7, 8, 9) has provided considerable evidence concerning the purposes served by the component parts of the whole system. An ability to maintain considerable rigidity of the trunk is associated with: long sectors, traversing more than one segment, of the dorsal longitudinal systems; well developed deep dorso-ventral and deep oblique muscles; and scanty superficial pleural muscles (Lithobiomorpha, Pauropoda). Considerable flexibility of body is associated with opposite characteristics: short sectors of the dorsal longitudinal muscles (Geophiomorpha): deep dorso-ventrals and deep obliques weakly developed, present on thorax only (Diplura) or absent (Symphyla); and superficial pleural muscles well developed either in great variety (Geophilomorpha, Scolopendromorpha and Diplura) or in size (Symphyla). Maki (193 8) figures short dorsal and ventral longitudinal muscles for the abdomen of the many pterygotes he describes. It has been noted above that Tomocerus and Orchesella maintain conspicuous rigidity of trunk, which is necessary for the use of hydrostatic forces in the jumping mechanism. It is not surprising therefore that the musculature of the Collembola shows: long sectors of the dorsal and sternal longitudinal system traversing several segments; very well-developed deep dorso-ventral and deep oblique muscles and the only superficial pleural muscles present are highly specialized and serve to maintain shape and rigidity and do not promote flexibility. The jumping Collembola show, in addition to the above, a very welldeveloped lateral longitudinal system such as characterises the Onychophora and epimorphic Chilopoda. These muscles are fused with the sternal longitudinals in the anamorphic Chilopoda and are absent in Pauropoda, Symphyla and other hexapod classes. It is significant that the Onychophora, epimorphic Chilopoda (Geophilomorpha and Craterostigmidae in particular) and the Collembola all employ changing haemocoelic pressures for their several
98 300 S. M. MANTON ways of life (Part 8 and above, 0 9A). The dorsal, lateral and sternal longitudinal muscles form a very substantial cylinder much as in the Onychophora and Geophilomorpha (figures and Part 8, figure 47), broken dorsally by the heart and ventrally by the nerve cord and structures above the ventral groove. Moreover the systems overlap, as shown on figure 27P, where the dorsal sectors are marked by letters, the lateral by numbers, and the sternal by letters. But the strength of the cylinder of muscles is probably enhanced by the complexity of superimposed long sectors, forming roughly superficial and deep layers, and these are made possible by the presence of the intersegmental tendon system giving sites of origin to both superficial and deep muscles (see below). The great development of the dorsal, lateral and sternal longitudinals, the deep obliques, and the many long sectors of the dorsal and sternal longitudinals must provide the pressure. The antagonists are the deep dorso-ventrals, much smaller in bulk, whose function is the elongation of the contracted longitudinals and deep obliques. The great length of abdominal segment 3 in some of the jumpers (e.g. Tornocerus), together with the large size and separation of the ventral tendons of origin of deep dorso-ventral muscles dv and deep oblique muscles ob (figure 21a), must be of particular importance in augmenting the slight body shortening produced by the dorsal, lateral and ventral longitudinal muscles. Further functions of the muscles are given in Appendix I. The superficial pleural muscles (figure 20a) are very well-developed and appear to resist any tendency for the body wall to bulge outwards under pressure from the contraction of the longitudinals and deep obliques. All along the body they hold in the more flexible ventral parts of the cuticle. They are particularly massive in the 3rd and 4th abdominal segments where flow towards the manubrium at the jump must be greatest, and they are assisted by the tendons rn and n which are devoid of muscles, figures 20a, 260, 27R. The details of the superficial pleural muscles and of those of the hamula and ventral tube are given in Appendix 1. The conclusions reached concerning the trunk muscles of a jumping collembolan are listed at the end of the descriptions of the components of each category in Appendix I. The comparisons between the trunk musculature of the jumping and non-jumping Collembola is given in 0 9B (iii) below. (v) The endoskeleton of Tomocerus The endoskeleton is entirely tendinous, and there are no apodemes or other cuticular intuckings, such as constitute the major part of the endoskeleton of the Diplura (figures 10-13). The endoskeleton comprises complexes of tendinous sheets, staining blue with Mallory s triple stain, which are essentially intersegmental, but elaborated in various ways on the thorax and abdomen. There are also tendon complexes which are segmentally arranged in association with the superficial pleural musculature; and there are large tendons elaborated to serve special purposes, such as the sheets rn and n supporting the ventro-lateral folds of the abdominal body wall, shown in black on figures Ribbon-like tendinous insertions. These insertions on the body-wall
99 EVOLUTION OF HEXAFOD LOCOMOTORY MECHANISMS 301 characterise the Collembola and have not been found so far in any other class. Groups of ectodermal cells of enormous size, with large nuclei and packed with tonofibrils, form either inwardly directed cones or flanges. From these wide tono-fibrillar bases arise the more compact tendons which take the form of rods, rope-like ties of various kinds, short or long tendons of origin of muscles or of endoskeletal plates, etc. The many smaller tendinous insertions on the thoracic body wall are of the same essential nature as the very large structures present in the abdomen (figures 20a, 21, 22 marked tonofibrils, 26N, 0,27P, R). The intersegmental tendons. These are seen most simply in the dorso-lateral intersegmental tendons of the dorsal and lateral longitudinal muscles. They arise at each intersegment, all along the body, from the anterior incurled margin of each tergo-pleural arch. They are shown typically at thoracic intersegments 2-3 and thoracic 3-abdominal 1 in figures 18a, 26 M. The thick incurled cuticle ( cuticle 3 on figure 26L) carries the superficial tendons, marked in figure 26M. In addition there is a deep tendon, marked on figure 26M, in the form of a sheet, attached below by a fan of tonofibrils to the body wall cuticle at the lower limit of the tergo-pleural arch, and above by another fan of tonofibrils to the dorsal cuticle just posterior to its intucked thickened anterior margin. The superficial tendinous sheet and its processes carry all sectors of the dorsal longitudinal muscles, for example muscle OC from abdominal segment 2 is ending in section L while muscles IE and OE originate in sections L and M and then pass posteriorly. This superficial tendinous sheet also carries some sectors of the lateral longitudinal system. Muscles 3 and 4 from segment 2 insert superficially just posterior to the level of figure 26L and are replaced by sectors 1 and 4 shown round the tendon in figure 26M. Muscle 2 in L is seen again in section M, ending on the inner face of the deep tendon (marked). Muscle 1 in section L is associating with tonofibrils (black) of the deep tendon seen in section M where muscles 1 from segment 2 is ending. That the tergo-pleural arch covering thoracic segments 1 and 2 is a fusion of the units appertaining to these two segments is shown by the presence of a typical intersegmental tendon attached to the intersegmental region of this common sclerite. This tendon, marked tendon e on figure 18a, carries typical lateral longitudinal and reduced dorsal longitudinal muscles. The ventro-lateral intersegmental tendon system is more diffuse. In the abdomen, ventral to the tergo-pleural arches, the intersegments are ill-defined and tendons insert on the body-wall both anterior and posterior to the intersegmental planes. In the thorax the intersegments ventrally are clearly defined, and the tendon system spreads forwards into the segment in front forming the major paired plates of endoskeleton g, bearing many anchors to the body-wall at and in front of the intersegmental line, and important processes pass backwards from these plates into each segment behind. The ventro-lateral tendon system gives the site of origin to the deep dorso-ventral dv and deep oblique ob muscles and to the sternal longitudinal muscles all along the body (figure 21). In the thorax a stout arm leaves each tendinous plate g on either side, and supports muscles dv and ob in the following segment, as shown in figure 18a. There is no plate g in the abdominal segments. On either side of the middle line, ventrally and behind abdomen 1-2
100 302 S. M MANTON intersegment, a stout sheet of enormous tonofibrils is attached to the cuticle. The enlarged ectodermal cells form an antero-posteriorly extended ridge, the contained tonofibrils contracting into a rope-like tendon which bears muscles dv,ob and the sternal longitudinals, as shown in figure 21a. At abdomen 2-3 intersegment the tendon of origin of muscles dv and ob is divided and the tonofibrils longitudinally spread out into two ribbons. The part carrying muscle ob lies in the posterior part of abdominal segment 2, figure 26M. The ribbon origin of the tendon carrying muscle dv and sectors of the sternal longitudinal muscles (figure 21a) lies in the anterior part of segment 3, figure 26N. All three ribbon tono-fibrillar plates just described on abdominal segments 2 and 3 have sites of origin on either side of the middle line at the sides of the median furrow. The homologue of the enormous ribbon tono-fibrillar plates and tendons of abdominal segments 2 and 3 appear to be tendon r in abdominal segment 4 (figures 21a, 27s). The tonofibrils are short, as is the stout tendinous mass between them and muscle dv, so enabling this muscle to pull directly on the highly sclerotized basal rod of the basal plate on either side. Owing to the folding-up movements of the springing organ and associated structures, there is no muscle ob in segment 4, and therefore no muscle for dv to elongate. Muscle dv appears to have changed its function, straightening the basal rods at the jump instead of contributing to the recovery movements. In abdominal segment 5 tendon 4, described above 9 9A(ii), is associated with the homologues of muscles dv and ob, and the presence of the springing organ must account for the position of origin of this tendon. The paired endoskeletal plates. The plates g form the major part of the thoracic endoskeleton. They have been figured by Carpentier (1949, fig. 5) and are shown here in figures 22-25A-J. They are represented diagrammatically in figures 18a and 21a because their confines are so complex in outline. The plates carry sectors of the sternal longitudinal and the deep dorso-ventral musculature. Antero-ventrally the plates lie mainly below the longitudinal muscles (figures 22-24A, D-E, G the lower g), and rise through these muscles to reach their dorsal extents (figures 22C, 24G the upper g, and figures 255 g). This most dorsal part of each plate gives the site of origin to the deep dorso-ventral and deep oblique muscles dv and ob (figures 18a, 22-25C, G, J). Anteriorly each plate forms a stout node of complex form situated over the middle of the coxal cavity and bears extrinsic suspensory leg muscles, a category not present in any arthropod so far considered (Parts 6-9, and needed here because there is no coxa-body articulation, (9 7B(iv)). This skeletal node is marked G on figure 18b and is clearly drawn, but unlabelled, on some thoracic segments in figure 18a. Since the pull by the muscles is exerted on the plates g in many directions, it is not surprising to find a system of anchors to the body wall which are much more complex than those of Diplura which possess a rigid cuticular base to the endoskeleton. These ties are present just where there is functional need for them, and they are not exactly homologous from one thoracic segment to another. The ties on the second and third thoracic segments are labelled by the letters used by Carpentier (1949), and the ties in the first thoracic segment are numbered 1-10 because most of them do not correspond with those of other thoracic segments. The prothoracic ties radiate forwards, laterally and

101 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 303 ventrally, as suits the functional needs, and differ considerably from the ties on the meso- and metathorax. There is a large measure of similarity between the ties on the meso- and metathoracic segments where the leg bases are set well apart on the sternal face of the body. The plates g are anchored by the processes k on either side which unite with thickened subcutaneous connective tissue (black line in figure 24H), from which a tonofibrilar system links the whole to the cuticle of the tergo-pleural arch. On the mesothoracic segment the elaborations of process k, with four separate roots to the cuticle (figure 23E), is occasioned by: the coxal suspensory muscles of leg 2 pulling directly downwards from process k (cf. muscles sus.3. and sus.4. on figure 23E and 24H); because the sternal longitudinal muscle kb (figure 21b, 23E) arises just anterior to section E and pulls directly on the tie k; and because the retractor coxal muscles ret.co.a. insert on this process on the mesothoracic segment, but on to the tergopleural arch on the metathoracic segment (figure 18a). Laterally the plates g on meso- and metathorax are linked by a bifid rod r to the proximal parts of the proximal coxal segment as shown in figures 23E, 24H. The rods appear to end compactly against the cuticle without a wide fanning of tonofibrils. On the metathoracic segment (figure 24H) a rodp leaves the basal part of tie k to end on the proximal margin of the pleurite. There is neither pleurite nor rod p on the mesothoracic segment, although drawn by Carpentier 1949, fig. 5. Ventrally the ties between the endoskeletal plates and the cuticle are numerous and are small editions of the ribbon tonofibrillar junctions noted above. They are described in detail in Appendix 11. There is considerable similarity between the ventral ties of the meso- and metathoracic paired endoskeletal plates, which differ from those of the fused prothoracic plates. The details of endoskeletal structure demonstrate a flexibility in form of the same general pattern. The similarities between meso- and metathorax, with minor differences correlated with function, appear to be due to needs rather than to phylogeny. There can be nothing fundamental about a pattern of ties of the endoskeleton if they can be so very different (see Appendix 11) on the prothorax compared with the meso- and metathorax of the same species. (vi) Cuticular features of the collem bolan trunk It has been shown above how the trunk musculature and cuticular elasticity provides the force which effects the propulsive stroke of the springing organ. This muscular force, delivered via the trunk haemocoel, is most economically employed when there is no outward bulging of the body wall or of the cuticle at the leg joints when the trunk muscles increase the internal hydrostatic pressure. Many features have already been noted which contribute towards this end; the limitation in the areas of flexible body wall; the absence of typical arthrodial membranes between the trunk segments; the absence of a coxa-body articulation, the leg being suspended firmly by an internal and unique suspensory system which holds other leg joints as well; the small number of leg segments, and therefore fewer distal joints with their flexible inter-podomere cuticle to be safeguarded against outward dilatation; other leg features, see 3 7B(i), (iv); the muscular and tendinous ties which prevent the flexible parts
102 304 S. M. MANTON of the body wall and ample coxa-trochanter arthrodial membrane from bulging outward (e.g. thoracic endoskeletal ties, trunk tendons m. and n. (figures 26,0, 27R), see 5 9A(v); and muscle a and some suspensory muscles from the coxa-trochanter arthrodial membrane (figures 23D, 24G). The remaining cuticular features which contribute towards the same need are considered below. The cuticle of the jumping Collembola is remarkable for the high degree of sclerotization everywhere and for the absence of typical arthrodial membranes. Figure 16 shows sections of cuticle of Tomocerus from a representative variety of regions, the conventions being the same as used in Parts 5, 8. The mechanical properties of myriapodan cuticles in relation to the structural differentiation shown by Mallory s triple stain, were considered in Part 5, p Arthrodial membranes of most arthropods are flexible and ample in area, except near the actual articulating or imbricating sclerotizations at a joint. Usually they are composed of a thick layer of cuticle staining pale blue with Mallory s triple stain, covered by a thin outer layer of amber coloured cuticle (Parts 5, text-fig. 8; 6, text-fig. 14; 7, text-figs. 3, 14, 10, 17-22; 8, figures 6, 11-15, 76). The highly sclerotized surface layer does not readily stretch, but it is thin enough to permit easy flexures of the membrane. The thick, less sclerotized cuticle staining blue with Mallory is stretchable and deformable in all directions. Tomocerus, in contrast to the myriapods illustrated by the figures noted for Parts 5-8, shows very short intersegmental connections between the trunk sclerites (figures 18a, 21a). This accounts for the marked trunk rigidity of the body at all times, cf. the different views of the body in P1. 3, figures a-f. The non-jumping Collembola have much more ample intersegmental arthrodial membranes and consequently greater mobility (PI. 3, figures h-i). The tergo-pleural arches of Tomocerus are well sclerotized and cover the dorsal and lateral longitudinal muscles, the latter being absent in other hexapods (figures 15d, 16, 22-28). The limits of the sclerites usually cannot be shown on the scale used in figures A clear anterior thickening of the cuticle of tergo-pleural arch 3 on the abdomen is seen on figure 26L, marked cuticle 3, this thickening fades out ventrally, as seen in the ventral half of both sides of this section (figure 26L, M). Apart from a thickened anterior margin to the trunk sclerites, they fade into the more flexible surrounding cuticle, but their limits are well shown by staining with chlorazol black (figure 28). It is suggested here that the major series of trunk sclerites are not simple tergites, as has hitherto been supposed, but that they are tergo-pleurites, a type of sclerite not present in the other apterygote classes, all lacking a lateral longitudinal muscular system. The presence of a small pleurite on the metathorax alone is not incompatible with the presence of a tergo-pleurite above it, since pleurites can be many. Diplopoda show some examples in which tergites and pleurites are separate, the lateral longitudinal muscles (retractor paratergalis, etc.) lying internal to the pleurites (e.g. Siphonophora, Part 7, text-figs. 9, 11, 13), but more often the tergal and pleural sclerotizations form one piece (Parts 4, text-fig. 3a; 7, text-figs. 21, 22, 24, 25, etc.). In he thysanuran Petrobius the thoracic and abdominal tergites are much prolonged into paratergal lobes reaching the sternal level, but these free lobes, composed

103 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 305 of closely applied inner and outer folds of cuticle and ectoderm (figure 32), as in a crustacean carapace, only superficially resemble the lateral parts of the tergo-pleural arches of Tomocerus. The tergo-pleurites in Tomocerus provide trunk rigidity by each sclerite enveloping much of the body surface. The arches of the first and second thoracic segments are fused in Tomocerus, the cuticular insertion of the longitudinal muscles marking the intersegmental boundary (figure 18a). This fusion has been noted by Carpentier (1949), showing that the pronotum is not absent. This is yet another feature contributing towards trunk stability, and stands in marked contrast to the features already considered which provide thoracic flexibility in the Diplura (3 8). Laterally above the legs the tergo-pleural arches tuck inwards, as shown in figures 22B, C, and then merge into the thinner cuticle around the leg bases. The edges of the thoracic sclerites are held firmly in place by the pleural muscles (figure 20a), the two sectors of pleural muscle thorax 2 (figures 18a, 22C, 23D) indirectly pulling on the middle of the ventral part of the fused pro- and mesothoracic sclerites. The metathoracic sclerite is strongly held by pleural muscle thorax 3 (figure 24H). The cuticle of the ventral surface of the body is thin and flexible except for the basal sclerite 4 below the fourth abdominal tergo-pleural arch (figure 15a, c). There are no typical sternites anywhere else. Mid-ventrally a cuticular gutter leads from the tip of the ventral tube to the labial gap into the pre-oral space (figures 22, 23, 24, 251, 161) and there are two median deep tubular depressions just in front of legs 2 and 3 from which the tonofibrillar connectives 10 and bm. pass to the endoskeletal plates in front (figures 18a, 22C, 23F). But, as already noted, the ventral cuticle is well tied to the endoskeleton by a very large number of tonofibrillar junctions (Appendix 11), and its shape is under control. Thus all the features concerning cuticular structure, the form of the sclerites, the associated muscles and endoskeletal ties to the cuticle, the small number of leg joints and their great specialization, are associated with the maintenance and control of haemocoelic pressure of the body used for jumping in Collembola. 9B. CONCLUSIONS CONCERNING COLLEMBOLAN STRUCTURE It has been shown in 3 9A(i), (vi) above how the cuticle of the entire body of Tomocerus, the proportions of the segments, and the trunk and limb musculature serves the abdominal jumping mechanism. The stabilizing extrinsic muscles of the jumping organ give only slight aid to the main propulsive force, produced by sudden increase in internal hydrostatic pressure, resulting from the actions of the trunk musculature, assisted by elasticity of the basal rods. The largest extrinsic muscles of the jumping organ are used to pull it back to its resting position. A summary of the functional significance of the details of muscles in each category is given at the ends of the descriptions in Appendix I. (il Cuticle and musculature A primary function of the cuticle is the manner in which it prevents unwanted billowing out of the more flexible regions under increase of internal
 is shown by the great difference between")
104 306 S. M. MANTON pressure. That the muscular component of the jumping mechanism receives only subsidiary assistance from the cuticular elasticity of the basal rods (figure 15a, c) is shown by the great difference between collembolan trunk muscles and the simpler series of muscles described for other hexapods (Snodgrass, 1929, 1931, Maki, 1938 etc.). The latter consist mainly of short muscles crossing one intersegment, and lack a lateral longitudinal system. Had cuticular elasticity been the primary source of energy causing the jump, the trunk muscles would be expected to be less unlike those of other hexapods. The elasticity of the cuticle of the manubrium is used, along with intrinsic muscles, in controlling the movements of the dentes. Most of the trunk muscles serve the facilitation of trunk rigidity and slight, but very strong, longitudinal contraction anterior to abdominal segment 4, increasing the general hydrostatic pressure which blows out the springing organ at a jump. The great length of abdominal segment 3, in contrast to the preceding and succeeding segments (figures 6m, 21), gives particular length to the deep oblique muscles and to the dorsal, lateral and sternal longitudinal muscles of this segment. It is the anterior ends of these muscles which are approximately fixed in position relative to their posterior insertions. The extra length of these muscles will therefore cause a larger displacement of tergo-pleural arch 4 and of the anterior-lateral edges of basal plate 4, hence increasing the pressure flow into the manubrium, than could shorter muscles contracting by the same proportion. The great length of abdominal segment 3 will therefore enhance the hydrostatic squeeze towards the mouth of the manubrium and also increase the efficiency of the single antagonistic pair of muscles 1 of the lateral longitudinal system, which fold up the manubrium after the jump. This heteronomy in segment lengths in the abdomen is absent in the non-jumping hypogastrurids, etc. but is not always as marked as in Tomocerus among other jumping species (e.g. Orchesella, pl. 3, figure f). Maki (1938) made a comparative survey of the musculature of the thoracic and anterior abdominal segments of 48 representative insects. This account, although lacking in accuracy and in detail, does show the presence and absence of certain salient features. He classes the lateral longitudinal muscles of Collembola as part of the dorsal system, giving no details. He shows a few long muscles, crossing two successive segments, in Lepisma and Folsomia, alone among hexapods; and he shows an abundance of short muscles, extending from one thoracic or abdominal segment to the next, in all other insects. Snodgrass (1931) in his survey of the abdominal musculature of apterygote and pterygote insects finds no true lateral longitudinal muscles. He differentiates a paratergal, or most lateral sector of the dorsal system, which he states is not commonly present, but is well developed in the Acridiidae, uniting the margins of successive tergites above the line of the spiracles, and is separated from the rest of the dorsal system by the insertions of the deep dorso-ventral and deep oblique muscles (his Musculi luterdes). Thus the Collembola differ strikingly from other hexapods in two major respects, (1) the presence of lateral longitudinal muscles and (2) the abundance of long sectors in the dorsal and sternal longitudinal systems. The lateral longitudinal system resembles that of the epimorphic Chilopoda (Part 8, figures 38, 41,47, 50, 51, 53) which is pleural in position and well below the spiracles. The onychophoran lateral longitudinal muscles are similarly situated.
105 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANlSMS 307 (ii) Endos kele ton The collembolan endoskeletal system, composed entirely of connective tissue fibres with no apodemes, is very different from that of the Diplura. Some general similarities exist between Collembola and other arthropods in, for example, the tendons of origin of deep dorso-ventral and oblique muscles, cf. thorax of Tomocerus and Japyx, but with some differences in Campodea, figures 18a, 21a, 10, 13a and see also Chilopoda in Part 8, figure 51 muscles dvmp.,dvma.,dvtr. These similarities are functional and do not mask the differences in plan of endoskeleton in Diplura and Collembola. The ribbon-tendon insertions of many muscles of Collembola are unique and are related to jumping. (iii) The structure of jumping and non-jumping Collembola It is useful to consider briefly the similarities and differences between the skeleto-muscular systems of jumping and non-jumping Collembola. It is probable that the Collembola which do not jump are secondarily so, although they show some features which are probably primitive: the separate pro- and mesothoracic tergo-pleural arches and the third abdominal segment not being unduly long. The hypogastrurid and onychiurid Collembola do not jump, and they display much greater flexibility of trunk as they turn in and out of small crevices, than is seen in the jumping Collembola (pl. 3, cf. h, i with a-f). The cuticle at the joints is more ample and less sclerotized than in the jumpers, so permitting greater trunk bending, which is also facilitated by the free prothoracic tergo-pleural arch and the even series in length of the segments. There are no ventro-lateral abdominal ridges, or associated tendons, because there is no springing organ. The musculature and endoskeleton of Onychiurus sinensis Stach. is basically as described here for Tomocerus. The three thoracic plates g are present, even if linked to the cuticle slightly differently. There are segmental ventro-lateral endoskeletal nodes along the abdomen, bearing the deep dorso-ventral, the deep oblique and the sternal longitudinal muscles. These nodes are joined transversely by fine strands staining blue with Mallory (absent in Tornocerus), and they lack the enormous tonofibrillar junctions to the cuticle. Junctions are present, but they are not conspicuous. The mobility of the onychiurid trunk is mediated by the free intersegmental joints and by muscular features which may well have been derived from those of jumping Collembola. The four dorsal longitudinal muscles traversing a typical segment of Tornocerus are present. The Lateral longitudinal system, absent in all other hexapod classes, is also present. But the dorsal, lateral and sternal longitudinal muscles are relatively much smaller than in Tomocerus, they do not form a muscular cylinder round the body, and the sectors are not superimposed. The superficial pleural muscles are very well developed, but pass from the ventro-lateral endoskeletal nodes upwards and outwards to the margin of the tergo-pleural arches, not arranged as in Tomocerus. The transverse linkage of the nodes and the positions of the pleural muscles may mediate flexibility of trunk. The large muscle 1 from the lateral longitudinal system of the jumping Collembola which flexes the manubrium (figure 17a), is
106 308 S. M. WTON recognizable in abdominal segment 3 of Onychiums, but it inserts posteriorly and below upon the linked endoskeletal nodes on either side, at the junction of the third and fourth abdominal segments. The muscle runs obliquely, in an opposite direction from muscle ob. and roughly resembles muscle dvma. of the Chilopoda (Part 8, figures 41,42, 50, 51 etc.). A muscle in this position is not present in other segments, and it is probable that this muscle is the metamorphosed muscle 1 of the degenerated springing organ (figures 17a, 20b, 26N, 0, 27), and, as for muscle dvrnu. of the Chilopoda, probably controls trunk rigidity. The extrinsic limb musculature and the suspensory system of the legs are essentially alike in the non-jumping Onychiurus and in Tomocerus. Thus the skeleto-muscular features of non-jumping onychiurids could have been derived from the Tornocems type on abandoning both a jumping habit and the extensive use of hydrostatic forces, in favour of small size, short legs, and improved flexibility of body, suited to moving about in minute spaces. But some divergence in habits probably preceded the present day refinements of the jumpers, and occurred before the fusion of pro- and mesothoracic tergites, or the full specialization of the third abdominal segment, took place. The jumping Collembola can therefore be taken as typical representatives of the class. The morphological evolution of the collembolan trunk appears to have been associated with jumping in a manner so fundamental and all embracing as to suggest the use of jumping escape reactions from the earliest differentiation of the class. Elsewhere among the arthropods jumping has been evolved many times, but usually as isolated cases in particular families and orders where the accomplishment does not involve the entire morphology. But thysanuran morphological evolution also has been associated with jumping movements by thoracic and abdominal limbs (see below). The small number of abdominal segments in Collembola, six only in both jumping and non-jumping species, is probably related to the basic jumping habit of the class. The abdomen acts as a hydrostatically firm unit whose intersegmental joints require rigid control. The dentes strike the ground roughly below the centre of gravity of the jumpers. If the abdomen was longer, as in the staphylinid larva on pl. 1, figure h, it would also have to be stiff and the springing organ would have to be much longer than is usual in order to exert a thrust below the centre of gravity. If the springing organ did not reach this level, somersaulting and no high jumping would result. The size of the springing organ is doubtless optimal for the forces employed by the hydrostatic mechanism and the latter is well suited by a six-segmented abdomen. No convincing evidence has been put forward indicating a secondary reduction in the number of abdominal segments in Collembola, and it may be assumed that the small number six is primitive and was established early along with the jumping habit.
107 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 309,, median furrow leg 3 Figure 15. (a-c) Orchesella villosa Geof. (d) Tomocems longicomis Muller. (a) Lateral view of the posterior end of the body with the springing organ extended, as at the end of a jump, drawn diagrammatically from a preparation of the cuticle alone to show the limits of the sclerites; the median furrow is not shown, see pl. 4, figure a. The posterior hinge between manubrium and body is marked by a black spot. The mobile anterior articulation between the arms of the basal rods and the manubrium is marked by a cross (see pl. 4, figure c). The arrow shows the direction of movement of this anterior articulation when the manubrium swings forwards to its resting position. (b) Lateral view of the posterior end of the body with the springing organ flexed forwards, almost to its resting position when the hamula impinges into the base of the furca (dentes). The point marked by a cross in (a) and the ends of the basal rods of the basal plate 4, are infolded and raised to the point marked by the cross in (b), corresponding with the arrow head in (a). (c) Ventral view of the abdomen with the manubrium directed posteriorly as in (a), and on a larger scale. The axis of the posterior articulation between the manubrium and the body is shown by the heavy dotted line. The anterior articulation, marked by a cross as in (a) and (b), is shown in detail. The stippled cuticle indicates the slots of lesser sclerotization in the proximal part of the manubrium (basal sclerite), ffigure 27U and figure 16N. (d) Diagram of the stereoscan micrograph shown in pl. 4, figure b to give the positions of the joints and the nature of the parts, ab the undivided coxa of leg 1 ; a and b the proximal and distal coxal segments of legs 2 and 3, pl. the pleurite of the 3rd thoracic segment, the tergo-pleural arches of the thorax and abdomen are numbered, those of the pro- and mesothoracic segments being fused into one sclerite.
108 310 S. M. MANTON
109 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 311 Figure 16. The cuticle of Tomocerus longicornis Muller. The outline above shows a transverse section across abdominal segment4, with the manubrium of the springing organ lying in the vend furrow along the basal plate 4 below tergo-pleural arch 4. Rectangles indicate the positions of sections of cuticle shown in (a-h). The thickness of the line representing the cuticle in this outline is not drawn to scale. The sections of cuticle (a-r) are drawn to the scale given above for 0.02 mm. On all sections the highly sclerotised surface layer of cuticle is orientated towards the top of the page. A key to the conventions is given below. The colours are those shown by average staining with Mallory's triple stain; manipulation of the reagents can give other colours; the series represents progressive degrees of sclerotization. (a) Through the mid-ventral cuticle, thin red layer over the dark blue; flexible but strong. (b) Through the thickened highly sclerotized rib on the basal plate on to which muscle dv pulls; very thick highly sclerotized outer part with thin under layer of red-blue. (c) Through the thinner flexible angle in the basal plate, amber coloured cuticle may be present but is inconspicuous above the red. (d) Through the thick dorso-lateral edge of the basal plate. (e) Through the thin flexible but highly sclerotized cuticle between the ventral plate and the tergo-pleural arch. (f) and (8) Through the lateral parts of tergo-pleural arch; thick, but less sclerotized than in (4. (h) Through the mid-dorsal cuticle. (i) Through the coxal cuticle of leg 1. (j) Through the pleuron above leg 1. (k) Through the arthrodial membrane at the joint between cox8 and trochanter. (I) Through the mid-ventral thoracic cuticle showing red-staining walls of the longitudinal gutter. (m) Through the basal plate 5 (see figure 15a). (n) Through the flexible joint in the base of the manubrium, the position is marked by a nng n on figure 27U. (0) Through the hinge line between tergo-pleural arch 5 (left) and basal plate 5 (right), for position see figure 15a, b. (p) Through the cuticle forming one side of the posterior furrow on the manubrium, the position is marked by a rectangle p on figure 17W. (r) Through the cuticle forming the lateral face of the manubrium, the position is marked by the rectangle r on figure 17W.
110
111 EVOLUTlON OF HEXAPOD LOCOMOTORY MECHANlSMS 313 Figure 17. The jumping organ of Tomocerus Iongicomis Muller. (a) Diagrammatic longitudinal view of the jumping organ with its extrinsic and intrinsic muscles, tendons shown in black. (b) Longitudinal section of the junction between manubrium and one dens, passing through the pore connecting the internal cavities of the two, at a higher magnification than in (a). (c) Longitudinal section passing through the origin of muscle 1 from the proximal part of the manubrium, the position, c, is ringed in figure 27T. Comparison with the sections of cuticle of other parts of the body in figure 16 shows the enormous thickness of the sclerotization (white) on the manubrium, conventions as on figure 16. The tonofibrils of muscle 1 only are indicated. (W>(V) are sections across the manubrium at the levels indicated by the labelled arrows on (a); the antero-lateral insertion of muscle sheet mb is so oblique that the tonofibrils are cross cut. The cuticle at p and r is shown at a higher magnification in figure 16p, r respectively. Section V shows the unlabelled notch at the base of the dentes into which the hamula fixes when the jumping organ is flexed forwards. Tendon a is cut just proximal to its insertion on the cuticle.
112 3 14 S. M MANTON
 Longitudinal reconstruction of the principal components of the musculature of the trunk and the proximal musculature of the limbs, in lateral view. The viscera are omitted.")
113 Figure 18. Tomocerus longicomis Muller. Thoracic musculature and endoskeleton. (a) Longitudinal reconstruction of the principal components of the musculature of the trunk and the proximal musculature of the limbs, in lateral view. The viscera are omitted. The coxae of all thoracic limbs are ventrally directed, but they and the telopods (trochanter and femur) are flexed laterally so that the femur of each becomes almost horizontal. The coxa of leg 1 is twisted on the body so that the telopod and claws are forwardly directed and the morphologically dorsal surface of the coxa lies at its left-hand margin, the view through the posterior face of the coxa being drawn in detail. The coxa and telopod of leg 3 are twisted on the body in the opposite direction, so that the claw is directed somewhat posteriorly and the anterior face of the coxa is obliquely shown. The telopod of leg 2 projects directly sideways and its coxa is shown in the morphologically dorsal aspect. The arrows show the directions of twisting on the body of coxae 1 and 3. The several sectors of the dorsal, lateral and sternal longitudinal muscles are not distinguished from one another because they are so superimposed in this (and all other) aspects. The details of these systems are given in summary in figures 19, 20, 21, and in the transverse sections on figures (see Appendix I). Endoskeletal tendons are shown in black, but the ventro-lateral tendons g in diagammatic manner, because of the complexity. The prothoracic endoskeletal 3 r plate g is drawn separately in (b). A stout endoskeletal node C lies over the coxal cavity. C The coxa of leg 1 is undivided and marked ab; the two coxal segments of leg 2 are marked a and h; and on leg 3 the coxa is incompletely divided into two, =] segments a and b being united on the median face of the coxa (figures 23, 24G, H, 251). The coxal tendinous endoskeletal plate is shown in white and marked coral 0 z plate in its several aspects on the three legs. Leg 1 shows the proximal tendinous junction at the ventral margin of the nochanter on the right, unlabelled, in black. Distal to it lies the dorso-ventral trochanter tendon, uniting the dorsal trochanter margin on the left with a fan of tonofibrils (marked) to the cuticle on the right. On leg 2 only the proximal trochanter tendon is drawn, and both tendons are shown on the trochanter of leg 3. On the thorax two mid-ventral cuticular intuckings lie just anterior to legs 2 and 3 (figures 22C, 23F) on to which are inserted ties 10 and brn from the endoskeletal plates g of pro- and mesothorax. The tie marked bm posterior to leg 3 does not reach the middle line and is situated beside the anterior part of the median ventral tube (figure 25J). The posterior part of the head endoskeleton and its muscular connections are better seen in Manton (1964) figures (b) Shows the prothoracic endoskeletal plate (black) and its muscular and cuticular connections. The ties 1-10 to the cuticle are described in Appendix ' U s 8 3 N.e
114 316 S. M. MANTON
115 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 317 Fipure 19. Tomocerus longicornis Muller. Diagrammatic reconstruction of the plan of the dorsal longitudinal muscle system. Thoracic and abdominal segments are numbered on the right and the middle line is dotted. Short muscles, crossing one intersegment (S), are shown on the left, the outer (0) and inner (more median) (I) series being numbered according to the segment. Long muscles crossing two intersegments, or clear derivatives therefrom, are shown on the right. An inner (I) and an outer (0) series are present, marked A-G from before backwards. For details see Text, figures 18, and Appendix I.
116 318 S. M. MANTON
117 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 319
118 3 20 S. M. MANTON
119 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 321
120 322 S. M. MANTON FcpUres Tommends longicomis Muller, Transverse sections, at the levels indicated by A-U in figures 20a and 21a, to show the principal components of the musculanre and endoskeleton of the trunk and of the proximal parts of the legs. The series is completed by sections V and W, on the same scale, in figure 17, the levels of the sections being marked on f ire 17a. Tendons and tendinous endoskeletal plates are shown in black. The entire coxa of leg 1 is marked ab. The two coxal segments of leg 2 are marked a and b, and the corresponding regions, incompletely divided on leg 3, are similarly marked a and b. Only some of the intrinsic and extrinsic muscles of the legs are drawn on each reconstruction, but all principal muscles are illustrated, in this series and in figure 18. Different levels are shown on the opposite sides of some drawings, as indicated by the large letters, with or without a heavy vertical dividing line drawn across the figure. Three successive levels are shown on the marked parts of the section on figure 27P, Q, R. Each segment number is marked in the cavity of the gut or below the proctodoeum. All sections are drawn as if very thick and viewed from in front, and they depict much more than can be seen in one 10 pm section. Muscles cut approximately in the transverse plane are stippled; those cut approximately longitudinally are lined; sectors of the dorsal IongiNdinal system are marked by letters, see figure 19; sectors of the lateral longitudinal system are marked by numbers, see figure 2%; sectors of the sternal longitudinal system are marked by letters, but in different combinations from those of the dorsal longitudinal system, see figure 21a, b; the superficial pleural muscles are mostly cut obliquely and are marked by interrupted lines and by the lettering shown in figure 20a. The large tonofibrillar junctions of the tendons inserting on to the cuticle are marked tonofibrils in places on fires 22, 23J, 27P-R and are unmarked elsewhere. For details of the ties between the endoskeletal system and the cuticle, marked by numbers and letters, see text and Appendix 11.

121 Figure 22. (A) Passes through the tapering anterior end of the trunk. On the left the heavy stippling represents the external surface of the tergo-pleural arch, pleuron and coxa of leg 1, which are visible behind the cut edge of cuticle, indicated by a double line. This line lies where the endoskeletal ties 1-3 insert on the cuticle, leaving it bulging between them. The deep infolding above is that of the edge of the tergo-pleural sclerite where its anterior margin is intucked across the flexible neck, see figure 18a. The oesophagus is cut in (A) and the opening out of the mid-gut in (B). Muscles.q the anterior part of the coxa are shown. The dotted arrow indicates the course of dorsal longitudinal muscle SI. Th. 1-2 in front of the level of this section, see figures 18a, 19. (B) Through the middle of coxa 1 lookinginto its posterior concavity; muscle dep.tr.1. IS cut in (A) and continues to the trochanter in (B). (Bb) Lies a little posterior to (B)to show tie 7 from the endoskeletal system to the cuticle. (C) Section just posterior to the coxa-body union of leg 1. The posterior origin of pleural muscle thorax 2 is shown in figure 23D.
122 D F
123 EVOLUTION OF HEXAPOD LOCOMOTOKY MECHANISMS 325 Figure 23. Tomocerus longicomis Muller. Musculature continued, see legend to figures 22-27, p (D) Section through the thorax 1-2 intersegment between these legs showing the anterior face of the coxa of leg 2, drawn as if transparent. (Dd) lies immediately posterior to level (D), where the coxal adductor muscle add. inserts upon the endoskeletal tendon (black), supported by tie f; cf. tie q just in front which lies at the anterior end of the endoskeletal plate. (E) Section through the middle of coxa2, showing the elaborate process k extending dorsally from the endoskeletal plate g to unite with the cuticle by large struts of tonofibrils. The anterior arm of the plate, supported by tie q, is shown by dotted lines. (F) Section through the posterior part of coxa 2 looking into its posterior concavity. Cut muscle sus.co.pl. is shown entire in (E).
124 3 26 S. M. MANTON Figure 24. Tomocerus longicornis Muller. Musculature continued, see legend to figures 22-27, p (G) Section through the thorax 2-3 intersegment between legs 2 and 3, showing the anterior face of coxa 3, drawn as if transparent. The postero-dital margin of region u of the coxa is shown by dotted lines, the two regions, u and b, are united, ~ b on, the median side. The lower g represents the metathoracic endoskeletal plate and the upper g is the posterior extension of the mesothoracic endoskeletal plate which carries the ventral longitudinal muscles Ka and G (see figure 21b). (Gg) Section just posterior to (G) showing the origin of tieffrom the ventral cuticle, cf. mesothorax, figure 23D d. (H) Section through the middle of coxa 3.
125 EVOLUTION OF HEXAFQD LOCOMOTORY MECHANISMS 327 I J Figure 25. Tomocerus ngicornis Muller. Musculature continued, see legend to figures 22-27, p (I) Section through the posterior part of coxa 3 looking into its posterior concavity. Tie bm extends posterior to the metathoracic endoskeletal plate as in (J). (J) The left-hand half of the section (J) represents the body just posterior to the origin of coxa 3 and the right half of the section (K) is a little posterior to this the ventral tube being extended.
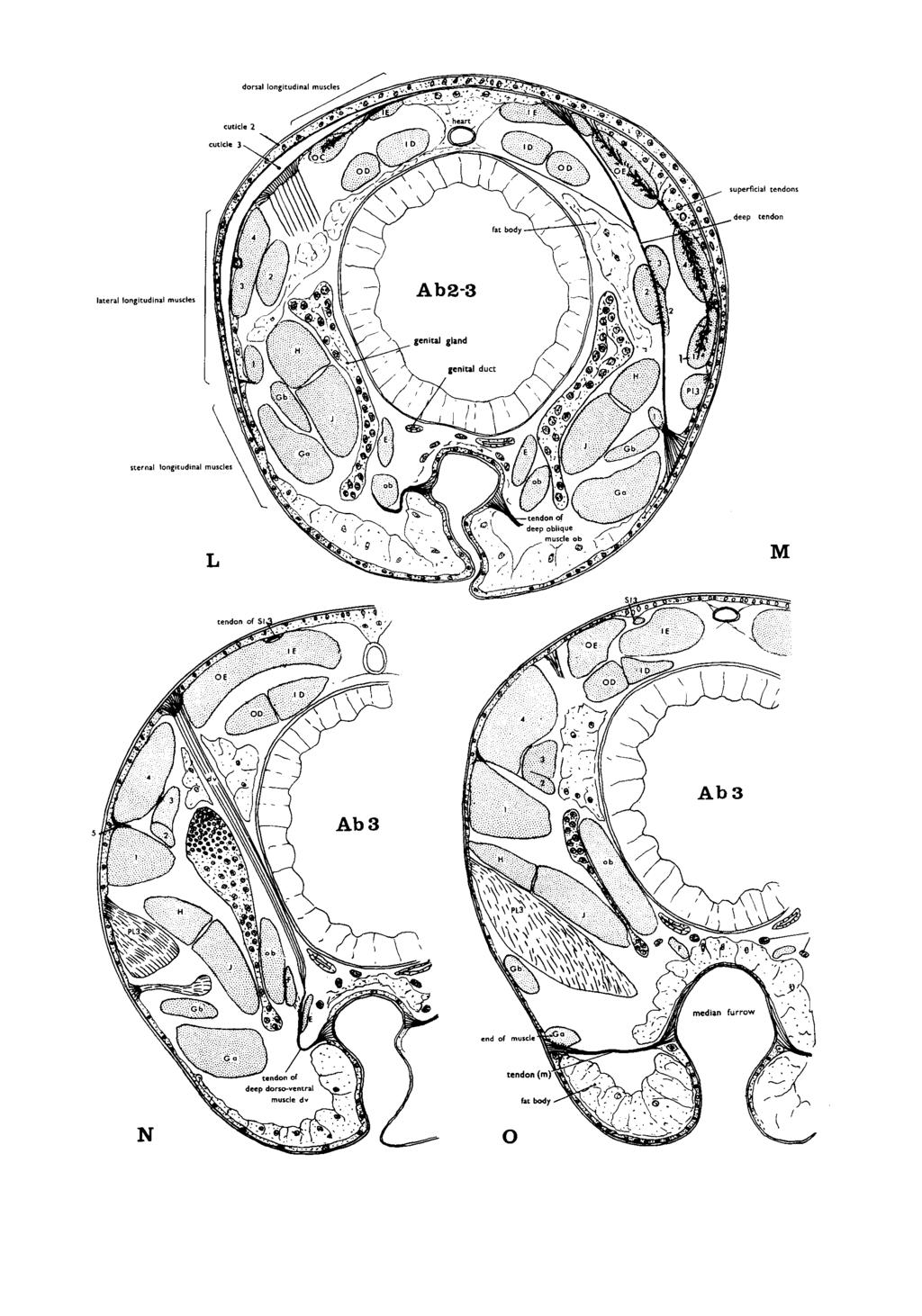
126 N
127 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 329 Figure 26. Tomocerus longicornis Muller. Musculature continued, see legend to figures 22-27, p (L-M) Section through the intersegmental region between abdominal segments 2 and 3, the level of (L) being anterior to that of (M). The dorsal overlap of the tergepleural arches is cut, the thick anterior margin being seen in (L) "cuticle 3". The origin of dorsal longitudinal muscle IE and OE from superficial tendons is seen in (M) while the lateral longitudinal muscles from abdominal segment 2 (sectors 14) are ending posteriorly on the deep tendon and on the superficial tendon complex. (N) Section through the anterior part of abdominal segment 3 posterior to level (M) to show the entire dorso-ventral muscle dv. and its tendon, the pleural muscle PL3. and the cylinder of dorsal, lateral and sternal longitudinal muscles. (0) Section through abdominal segment 3 anterior to the hamula to show the tendon m., the insertion of sternal longitudinal muscle Gu and the positions of the other major trunk muscles.

128 3 30 S. M. MANTON P Ikerrl longltudlnrl T U
129 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 331 Figure 27. Tomocerus longkornis Muller. Musculature continued, see legend to figures 22-27, p (P, Q, R) Sections through the 3rd abdominal segment, (P) at the level of the hamula to show its muscles and tendons and the large superficial pleural muscle P1.3. with part of its lateral insertion, (Q) lies a little posterior to (P) and shows the origin of the long muscle from the hamula tendinous junction to the anterior margin of tergo-pleural arch 4, as shown in figure 17a, and (R) slightly more posterior in level, shows another muscle from the ventral groove to the hamula complex at the origin of tendon n; the lateral end of this tendon spans the insertion of sternal longitudinal muscle Gb. The ventral origin of pleural muscle P2.3. lies in a groove lateral to the ridge bounding the ventral groove, further anteriorly this groove comes to lie on the median side of this ridge, as seen in (P), unlabelled. (S) Section through abdominal segment 4 cutting the basal rod on the folded basal plate and the tendon r., giving origin to dorso-ventral muscle dv. The genital duct lies below the proctodoeum. (T) Section through the anterior part of the base of the manubrium, the origin of muscle 1 is ringed at c, and shown at a higher magnification in figure 17c, muscle 1 (part of the lateral longitudinal system of abdominal segment 3) inserts on the anterior margin of tergo-pleural arch 3, as shown on figures 17a, 20b. (U) Passes through the hinge-like posterior articulation between the rnanubrium and abdominal segment 5, shown also by a black spot on figure 17a, b and a dotted line on c. The zones marked f are little sclerotized, see the stippled areas at the base of the rnanuhrium in figure 17c, the ringed cuticle at n is shown in figure 16n.
130 332 S. M. MANTON Figure 28. Orchewlla villoso Geof. Preparation of the cuticle of the anterior pan of the body, after treatment with potash and staining with chlorazol black. The more sclerotized parts of the cuticle appear grey, in contrast to the less sclerotized parts which remain clear, with the degree of staining employed. The cuticle is tom on the animal's right-hand side, so that it is opened out ventrally to show the middle line, the ventral tube and right leg 3. The corresponding parts of the coxa of legs 1-3 are marked a-b. The coxa of leg 1 is undivided, that of leg 2 is completely divided into two segments separated by short arthrodial membrane, and on leg 3 the two segments are united on the median side of the coxa. One pleurite only is present, situated on the metathorax. A certain amount of stretching of the less xlerotized areas has taken place. The disposition of the sclerites, here stretched apart, can be compared with the normal positions as shown by the stereoscan micrograph of Tomocerus longicomis, pl. 4, figure b.
131 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 333 trochanirr rendon a Figure 29. Tomocerus Iongicomb Muller, the leg in anterior view. (a) Normal leg 2 with the coxa-trochanter joint strongly flexed. The anterior components of the pivot articulations at either end of the trochanter are marked; muscle rer.fe., passes to the posterior component of the pivot joint. (b) Abnormal leg in which the trochanter-femur joint is absent and muscles dep.fe., zeu.fe., and retfe., are also absent. (c) Abnormal leg in which the nochanter-femur joint is absent, but muscles dep.fe., lev.fe., and *ef.fe., are present.
132 3 34 S. M. MANTON : anterior
133 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 335 Figure 30. Diagram to show the suspensory system of the collembolan leg. Left leg 2 is pendant from the body and viewed towards the side of the trunk. The coxa is approximately vertical and divided into the two segments a and b. The trochanter and femur are in line with one another and strongly flexed on the coxa, so that they project horizontally; they are foreshortened and viewed slightly from above. The tarsus is almost vertical and strongly flexed on the femur. The pair of small black spots at the distal end of the coxa marks the two components, anterior and posterior, of the coxa-nochanter pivot joint. The pair of small black spots at the distal end of the trochanter similarly marks the anterior and posterior components of the trochanter-femur pivot joint, but the right-hand spot lying on the posterior face of the leg is drawn as if the femur was transparent, and similarly for the distal part of muscle retfe. The heavy line drawn through each pair of spots marks the axis of movement. The single small spot at the distal end of the femur marks the dorsal femur-tarsus hinge articulation. Endoskeletal structures are shown diagrammatically in black. The large and complex endoskeletal plate forms the node G above the coxal cavity (see figures 18, 22-25). The tendinous process k suspends the node G from the tergo-pleural sclerite above and the endoskeletal plates are anchored in many directions to the cuticle. Muscles comprising the suspensory system are shown by double lines and muscles causing normal leg movements are omitted. The open double lines show some of the coxal suspensory muscles inserting on the node G; sus.co., sus.co.pl., and there are others. Muscles sus.co.pl. leave the antero-posteriorly placed coxal plate; and tendinous strands, unlabelled, leave the plate to end on the anterior components of the coxa-trochanter and trochanter-femur pivot articulations and the trochanter tendon. The latter spans the anterior part of the trochanter and is better seen in figures 22B, 23E, 24G, 29a, so leaving unimpeded central space for the femoral levator and depressor muscles. Suspensory muscles, marked by spots between the double lines, hold the trochanter tendon to the coxa. Hatched double lines mark suspensory muscles holding the posterior components of the pivot joint articulations: ret. tr. and rer. fe., and muscle ret. tn. holding the tarsus at its hinge joint with the femur. These muscles, termed retractor because of partial similarity with muscles of other arthropods, cause no flexures but prevent dislocation or separation of the articulations during moments of increased internal hydrostatic pressure.
134 336 S. M. MANTON 9C. THE MACHILID JUMPING MECHANISM The evolution of the trunk morphology of the Thysanura Machilidae must have been almost as extensively bound up with performing jumping gaits and airborne jumping escape reactions as has that of the Collembola (5 9B). High jumping is used by the Machilidae, but not by the Lepismatidae; however it is possible that some trunk features of the Lepismatidae may have arisen in association with jumping leg movements in somewhat the same manner as in the Machilidae. (i) Jumping Information on the mechanism of the escape jumps of Petrobius has been obtained from a study of anatomy and of tracks. The marks left on smoked paper by a series of, or by single, airborne jumps are shown in figure 37, printed in reverse Regions A and B are sites of landing from a previous high jump and of take off for the next. From landing site C the animal ran off by an irregular performance of the normal gait. At the take off of a high jump, marks are left on the smoked paper by the abdominal coxo-sternal plates, styles and often by the lateral margins of the abdominal tergites. The caudal filament and terminal cerci sometimes touch the paper and leave marks. No marks by the abdominal coxal plates are made on landing and marks are always recorded at sites of take off. The animals presumably land on their thoracic feet, the abdomen then being off the ground. The double lines are made by the tarsal claws being dragged along the paper. Figure 37b-e show the marks left at the take off of high jumps on a larger scale, the length of the animal, 10 mm, being shown by the vertical line. The marks formed by the coxal plates are not smudges, as would be left by organs being dragged over the surface of the paper. Smudges left by the horizontal movement of the sternal surface of a chilopod on smoked paper are shown in Part 3, text-figs. Sc, d, lob, and by the abdominal styli in normal running here in pl. 5, figure a. The precise marks left by the abdominal coxo-sternal plates indicate pressure exerted by these organs on the ground causing the jump. The distance apart of the extreme postero-lateral ends of the claw marks of legs 3 is that found in resting legs and in legs performing the normal gait. The legs 3 claw marks converge in the forward direction to a level which was in front of the head when the abdominal coxo-sternal plates were in contact with the ground. This means that the pair of legs was straightening by depressor and flexor movements, pressing against the ground as the animal was rising into the air. The records of figure 37d, e show one of each pair of tarsal claws ceasing to make a mark in the anterior direction before the other, and the final marks become intermittent anteriorly. These features indicate contact with the ground by the outstretched legs at the start of the jump, and that as the animal became airborne, the limb tips were dragged over the surface of the paper, the telopod straightening and adducting until these legs too left the ground. The irregularity of the claw marks anterior to those of legs 3, and their number, is due to both landing and to foot shuffling before the take off. The irregularity and variability of these marks do not suggest that legs 1 and 2 play more than a minor role in the take off of a high jump. Thus the soot paper records show
135 EVOLUTION OF HEXAPOD MCOMOTORY MECHANISMS that the abdomen is the main jumping organ, and that thoracic legs 3 may be of importance in keeping the animal on an even keel at the moment of the take off. 337 (ii) Cuticular features of the abdomen of Petrobius Not one of the many photographs and films taken of Petrobius show any lateral bending of the abdomen, but pl. 6, figures a-d show different degrees of flexure in the vertical plane giving a dorsal concavity. The abdominal tergites readily slide over one another dorsally, so permitting easy dorsal shortening. These tergites are remarkable for their paratergal extension to the level of the sternites (figure 38a, b) and for the enormous overlap of one by another (figure 3% d). Each coxo-sternal plate extends freely below the following segment. The anterior triangular area of each plate is usually considered to be the sternite while the rest is formed by flattened fused coxae, the styles representing their telopods. The cuticular junction between the sternal and coxal regions is shown in transverse section as a fold (figure 38e) which doubtless has the mechanical function of stiffening the anterior part of the plate where it is subject to muscular tension (see below). The antero-ventral face of the plate is well sclerotized from its anterior margin E to its posterior margin bearing the coxal sacs and styles (figure 3 8d). The postero-dorsal face is only half the length of the ventral face. From the posterior margin to F the dorsal cutkle of the plate is less sclerotized than the ventral face (figure 38c, d), and between the levels E and F on figure 38d the cavity of the plate is open to the haemocoel. A dorsal contraction of the abdomen, by a shuffling together of the tergites, is accompanied by considerable ventral expansion. The ventral surface between abdominal segments 1 and 5 on pl. 6, figures a, b is expanded by about 13% in figure c. A more extreme dorsal contraction probably takes place at an airborne jump, which must give a considerable ventral spreading of the coxal plates, and if the contraction of the muscles producing the dorsal shortening is very strong, an increase in internal hydrostatic pressure must occur momentarily, and may give a thrust against each coxo-sternal plate in the direction of the arrow on figure 38d. The overall tergal support by the overlapping lateral paratergal lobes from the tergites, and the way in which the overlaps support the anterior end of each coxo-sternal plate (figure 38c), must contribute to this end. Moreover strong muscles are attached to the anterior end of the coxo-sternal plate, whose contraction could both hold the anterior end firmly in place against any increase in internal pressure, and could tip the anterior margin inwards and upwards, so tilting the whole plate, thus depressing the propulsive posterior margin. These movements, executed speedily and with strength, and almost or quite synchronously all along the abdomen, must provide the force responsible for the high jumps by thrusting the posterior edge of each plate against the ground. (iii) The abdominal endoskeleton of Petrobius The endoskeletal structures of the abdomen of apterygote classes are usually less elaborate than those of the thorax. In the dipluran abdomen simple ventral
136 338 S. M. MANTON longitudinal muscles attach to the intersegmental marginal regions of the sternites and to thin ventro-lateral tendinous junctions, anchored to the intersegmental cuticle. In a japygid they appear much as drawn for the thorax of Gzmpodea in figure 13a, where they give origin also to muscle dvmd. But in the jumping Collembola (5 9C(iii)) and Machilidae the abdominal tendon system is well developed, and the form and complexities are correlated with jumping mechanisms and muscular needs. In Petrobius the thoracic endoskeletal tendon system forms elaborate transverse ventro-lateral structures situated largely near the intersegment and extending both forwards into the segment in front and backwards into that behind (see Barlet, 1967, Fig. 1 for Lepismachih). In the abdomen the endoskeletal tendons form a segmental pair of ventro-lateral systems separated mid-ventrally. The lateral part of each, marked lateral endoskeletal tendons, in figures 38d, 39 right, is extensive and diffuse, thin sheets of connective tissue fibres forming sites of origin and insertion for the deep oblique and other muscles. The most ventral part of the tendon system, marked coxal tendon on the same figures, extends forwards into the preceding segment and lies over the coxal cavity, supporting the coxa by muscle 19; the coxal tendon is compact and carries the sub-median sternal longitudinal muscles 15 on either face. The coxal tendon in the abdominal segments corresponds with the forward projection g over the coxal cavity in the thorax, see Barlet, 1967, Fig. 1. Fine strands of connective tissue pass from the main ventro-lateral complex to the ventral intersegment and to the anterior margin of the coxo-sternal plate (drawn on figure 38d, segment V). Dorso-laterally there are tendinous junctions between the segmental sectors of the dorsal longitudinal muscles 1-5, The tendons attach to the tergites well behind their anterior margins and sink deep into the body, in support of these massive dorsal longitudinal muscles, and send flanges to the insertions of dorso-ventral muscles 20, 21 (figure 39, segment V). It is probable that the dorso-lateral tendons also maintain a link with the major ventro-lateral system. (iv) The abdominal musculature of Petrobius The abdominal muscles of the Machilidae are some of the most remarkable in the animal kingdom. The description below and figure 39 represents the work of Dr G. G. E. Scudder which he has kindly allowed to be incorporated into the present account. Matsuda (1957) gave a short account of the muscles of Neomachilis, but the details are not confirmed below. However he suggests that this unique set up of the abdominal musculature is directly responsible for the characteristic strong leaping action, a conclusion with which we are in agreement, although differing profoundly as to the morphology. Petrobius shows some twisted ropelike muscles and others which loop round one another. Such muscles are recorded among arthropods only in the Malacostraca of caridoid facies capable of jumping backwards through the water by a strong flap of the abdomen e.g. Pdinurus (Parker & Rich, 1893), Homarus (Daniel, 1931), Praunus (Daniel, 1928, 193 l), Paranaspides (Daniel, 1932), Lophogaster and Gnathophausia (Daniel, ) Meganyctiphanes (Daniel, 1929) and see comparative musculature of these crustaceans (Daniel, 1932). Slightly twisted dorsal muscles occur in Geophilomorpha, Part 8,
 An inner, mesial series: muscles 1-4 arise from tendons near the anterior margins of the tergites and insert similarly on the following segment; Muscle 5 spans two segments.")
137 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS figures 42, 43. The dorsal longitudinal system consists of four series of muscles (figures 38d, 39). (1) An inner, mesial series: muscles 1-4 arise from tendons near the anterior margins of the tergites and insert similarly on the following segment; Muscle 5 spans two segments. All five muscles are twisted round one another like strands of a rope. (2) An outer, more lateral, series, of similarly twisted short muscles alongside muscles 1-5. (3) The dorso-lateral longitudinals comprise muscles 12, 13, situated lateral to the tergal insertion of the deep dorso-ventrals 20, 21. Muscles 12, 13 are untwisted, and extend from tendinous junctions behind the anterior margin of the tergite in each segment. (4) The oblique muscles 6, 7 arise antero-dorsally on the tergal face and extend postero-ventrally to insert upon the anterior margin of the following tergite. The ventral longitudinal system is simple and consists of two segmental muscles. The most median is muscle 15 linking the successive coxal tendons. It is not a bulky muscle and corresponds with the simplest ventral longitudinal muscles of other hexapods. Muscle 14 is more bulky and more lateral in position, and links successive ventro-lateral tendon systems. The deep dorso-ventral and deep oblique series of muscles are exceptionally well developed and elaborate. The deep dorso-ventrals are two in number. From its own tendon, anchored on the coxal tendon uniting ventral longitudinal muscles 15, muscle 20 extends upwards to insert on the tergite dorsally near its anterior margin, passing through the dorsal longitudinal muscles, as occurs in the thorax of Campodea, figure 13a. An extra dorso-ventral, musc2e 21, is present, arising more laterally from the ventro-lateral tendon complex; it inserts on the tergite immediately posterior to muscle 20. The deep oblique muscles are very bulky, with many additional sectors, and they traverse several segments. Muscle 22 represents the normal deep oblique muscle corresponding with ob. in Collembola (figure 21a) and duma. in Chilopoda (Part 8, figures 50, 51, 56), but is not over large. It is the deepest of the oblique muscles, arising from the ventro-lateral tendon complex and passing upwards and backwards to the anterior margin of the following tergite. Muscle 23 arises from the ventral part of the lateral tendon complex, and passes obliquely forwards, traversing the greater part of two segments, to insert on the lateral part of the tergite opposite the insertion of muscle 25B. This muscle roughly corresponds with the chilopod muscle dvmp. although traversing an additional segment. Muscle 24 arises from the anterior side of the ventro-lateral tendon complex and passes directly upwards and then backwards to insert on the following tergite, just ventral to the insertion of muscle 26. Muscle 26 follows a roughly similar course but traverses one more segment; arising from the posterior side of the lateral tendon complex, it rises to lie dorsal to and parallel with muscles 24 from the following segement, the two muscles insert together below the dorsal longitudinal muscles on the anterior part of the third and second segment respectively from their origin, as shown, figure 39. Muscle 25 is very bulky and forms two sectors A and B, arising from the mesial side of the ventro-lateral tendon complex. Both extend postero-dorsally to loop round muscles 24 and 26, passing external to them. Sector 25A then traverses the next posterior segment to insert on the anterior side of the tendon complex posterior to it. Sector 25B descends to the face of the following tergite, inserting opposite muscle 23 (figures 38d, 39). 339
138 340 S. M. MANTON Muscles from the coxo-sternal plate. All muscles from this plate arise from the parts usually considered to be coxal and are presumably derivatives of coxal extrinsic muscles. Muscles 27, 28, 29, 31 arise from the coxal plate anterior to the level of the coxal tendon. Muscules 27,28 from the edge and muscle 29, posterior to them, from the lateral face of the coxa, pass upwards and outwards to the tergite as shown in figures 38a, b. Muscle 31 arises near the mesial edge of the coxa, just lateral to the groove demarcating coxa and sternite, and passes upwards, mesial to the coxal tendon and muscle 19, to insert on the tendon of origin of muscle 20 (fwre 38a). Muscle 19 forms a short fan of muscles linking the coxal plate to the coxal tendon above it. Muscle 30 arises from the anterior side of the style base and passes forwards through the plate to insert on the anterior part of the tergite. 9D. CONCLUSIONS CONCERNING THE MODE OF ACTION OF THE JUMPING MECHANISM OF PETROBIUS While the jumping gaits of Petrobius are performed by the thoracic legs, the high jumping escape reactions are due to dorsal contraction of the abdominal segments, a thrust being exerted against the ground by the posterior margins of the coxo-sternal plates. The abdominal musculature provides strong and rapid increase in dorsal concavity of the abdominal tergites, with a corresponding ventral expansion, and a depression of the free margins of the coxo-sternal plates. The musculature also maintains strong antero-posterior rigidity, preventing any lateral flexure of the abdomen. The complexity and bulk of the abdominal dorsal longitudinal muscles is striking compared with those of other hexapods. The component muscles traverse one segment only, except for muscle 5. This arrangement resembles in principle the short stout muscles of Geophilomorpha, and, as suggested in Part 8, is suitable for causing strong longitudinal shortening of the body. The Collembola also need strong body shortening, but not localised dorsally, and their dorsal musculature (figure 19) is very different in detail from that of Petrobius, the two classes having evolved in independence (see 3 10G). The substantial and rope-like twisting of muscles 1-5, 8-11 in Petrobius resembles the twisted dorsal malacostracan muscles of jumping species (see above). The mode of action is unknown, but whatever it may be, extra length of each separate muscle is thereby obtained, compared with an unnvisted muscle, and this means that a smaller proportional contraction by the muscle can produce the same absolute shortening of the inter-tergite distance. Any approach to an isometric contraction is a physiological advantage. The whole mass of twisted muscle must be capable of very strong pulling together of the tergites dorsally. Constant alignment between the abdominal segments is maintained by dorso-lateral muscles 12, 13 and by the well developed dorsal obliques 6, 7. The latter correspond with the epimorphic chilopodan muscles s. ob, b. serving the Same purpose (Part 8, p. 273,Table 1, p. 362, figures41,43a, 55,56c, 57b). The deep oblique muscles, arising from the ventro-lateral tendons, are remarkable for their complexity (figure 39). The normal deep oblique muscle (see ob. Collembola, figure 21a) is present as muscle 22, but it is not enlarged as in the jumping Collembola in abdominal segment 3. An additional five deep
.")
139 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 341 oblique muscles, 23, 24, 25A, 25B, 26 link the tendon of origin with: tergite n + I by muscles 24, 25B; tergite n + 2 by muscle 26; tergite n - 2 by muscle 23; and ventro-lateral tendon n + 2 by muscle 25A (figure 38d shows the positions of the muscles diagrammatically). A pull exerted over a range of five tergites from each ventro-lateral tendon system cannot do other than strongly pull the tergites together dorsally. But the looping of the two muscles 25A, 25B round muscles 24 and 26 can presumably cause extra pulling together of the dorsal parts of the tergites. The great length of muscle 25A and the bulkiness of both presumably can deform muscles 24 and 26, so providing indirect shortening of the distance between their origins and insertions. Thus the looping probably increases the muscular force available for pulling the tergites together dorsally. The duplication of the normal deep dorso-ventral muscle 20 by muscle 21, arising more laterally from the ventro-lateral tendon, must provide additional antagonistic muscular force used to straighten the abdomen and pull out the contracted deep obliques. In the jumping Collembola dv., the homologue of muscle 20, is enlarged in the abdomen, but not duplicated (figure 21a). The ventral longitudinal muscles, in two main sectors, the smaller muscle 15 stabilising the coxal tendons and the suspensory coxal muscle 19, and the larger muscle 14 uniting the ventro-lateral tendon systems, are placed just where they are needed. Muscle 14 in particular must straighten the body after sudden contraction of the dorsal longitudinal and deep oblique muscles, but neither muscles will be doing conspicuous outside work, hence the simplicity of the ventral longitudinal system. The epimorphic Chilopoda show greater complexity of the sternal longitudinal muscles which are used in strongly shortening the body, and these centipedes also show complex arrangements of deep oblique and deep dorso-ventral muscles. The details differ from those of the Machilidae, but the functional assets are comparable. The musculature of Thysanura and Chilopoda have evolved in parallel, not one from another. The coxo-sternal plates must be tilted by contraction of muscles 27-29, 31 pulling on their anterior parts. A downward movement of the free posterior edge, and erection of the style by muscle 30, could assist the jump since the posterior edges of these plates form the site of take off. It is also possible that haemocoelic pressure in the direction of the arrow on figure 38d might assist the depression of the distal part of each plate. Strong contraction of the dorsal rope-like muscles and of the deep oblique complex might provide this pressure, the ventral longitudinal muscles being relaxed and extended. Thus it is concluded that the motive force for the high jumping escape reactions is provided by: the dorsal longitudinals 1-5, 8-11, the deep obliques 22, 23, 24, 25A, 25B, 26, and the extrinsic coxal muscles 27-29, 31 aided by muscle 30, an extrinsic from the telopod, which erects the style. The recovery movement, extending the dorsally contracted abdominal trunk segments, is effected by the deep dorso-ventral muscles 20, 21 and the ventral longitudinal muscles 14, 15. The maintenance of alignment is controlled by muscles 6, 7, 12, 13. The complete absence of superficial dorso-ventral and oblique muscles in Petrobius abdomen is in keeping with the conclusions already reached as to the function of these muscles (Parts 8, 9). They assist lateral bending of the trunk. They are well developed in the thorax of Diplura, in the epimorphic Chilopoda,
140 342 S. M. MANTON and in Symphyla, and they are entirely absent in the rigid-bodied Pauropoda, where well developed deep dorso-ventrals and deep obliques maintain rigidity, as in Collembola and Machilidae. Superficial pleural muscles in Collembola represent the superficial dorso-ventral and oblique category, here diverted to serve other purposes. Some jumping arthropods have mechanisms whereby the jumping organs are held in place while muscular tension is built up, and then released. The collembolan harnula is one such device used in holding the springing organ until its sudden release. Jumping dipteran larvae, such as cheese skippers (Sepsidae) and various fruit fly larvae e.g. Dacus (Olroyd, 1964), arch the body until the mandibles grasp the caudal end, a papilla easing the hold, and when the tension on the muscles is built up, the jaws let go, and the animal straightens and jumps, like a spring unbending. Neither Petrobius nor the Malacostraca have any external mechanism for holding either the jumping organs or the muscles while tension builds up, and it might be that looping and twisting of muscles has something to do with this need. Whether elasticity in some form is a special component of this muscular mechanism is unknown. Figcrre 31. Petrobius brevistylis Carpenter leg str~cmre. (a) Posterior face of left leg 2 to show the muscles and joints. More of the extrinsic muscles are seen in figure 32. The pivot pits (dicondylic, one pair of articulations on either side) between coxa and trochanter and between femur and tibia, are marked by white and by black spots respectively. Neither joint is equatorial, they are situated towards the ventral and the dorsal side of the leg respectively, giving great range of movement on the opposite side. The dorsal cuticle at the hinge between tibia and tarsus (white) is very stout and strong (6. Myriapoda, Parts 6 and 7). (b) Lateral view of the thorax to show the positions of the pleurites, the large overhang by the paratergal lobes, and the points of closest union of coxa and pleurite are marked by black spots. The coxal and pleural apodemes passing directly inwards in the transverse plane are not shown, see figure 32A and D. The levels A-G marked alongside are those of the transverse sections shown in figure 32.
141 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 343 PETROBIUS dorsal a

142 344 PETROMUS F
143 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 345 Figure 32. Petrobius brevistylis Carpenter. Thick transverse sections, viewed from in front: (A) through the prothoracic coxa and (B-G) through the mesothoracic coxa and associated structures, at the levels marked on figure 31b, (B, E ). under higher magnification than the rest. The coxal cavity is marked by intempted ruling; the conventions on B and E, denoting different degrees of cuticular sclerotization, are those given in the key on figure 16. On the other figures the cuticle which is conspicuously sclerotized is indicated by uniform black. The muscle numbers are those of Barlet (1967) for Lepismuchilis Y-signutu, but his muscle 117 is here shown to be tonofibrillar and not muscular, The specimen on sectioning became longitudinally shrunk and the intersegmental zones are pushed forwards into the segment in front in an unnatural manner. Level A passes through the coxal apodeme of leg 1 (there is no pleural apodeme, as on the meso- and metathorax) and through the two longitudinal pleurites marked X and Y. The anterior dorsal part of tergite 2 is cut, with its very strong and slightly mobile lateral junction with tergite 1, the latter extending ventrally as the paratergal lobe. The paratergal lobe of the mesothorax extends forwards and is just cut in the section, see lateral view in figure 31b for positions of the parts. The dorsal dlm. and ventral ulm. longitudinal muscles are marked. Level B passes immediately anterior to the sclerotized edge of the single mesothoracic pleurite, see level C. A thick cone of tonofibrils pulls on the pleurite and terminates above on the basement membrane below the tergal ectoderm. The paratergal lobe is formed by an outpushing from the tergite and its internal ectoderm is therefore double layered, see level D. Level C shows the anterior coxal articulation with the pleurite; the stout pleural protractor muscles 113, 114, which pass forwards to the tergite dorsally; the coxal adductor muscles 91,92 passing postero-ventrally, their cwal origins being seen in levels D and E; the coxal protractor, 101, arises from the anterior external margin of the coxa posterior to level C and from the tergal insertion of coxal abductor 102 this muscle passes posteriorly, through levels D and E, to its origin on flexible membrane just dorsal to the coxal margin in level F. This muscle may be partly protractor in effects. The pleurite is flexibly united with the sclerite Su on the underside of the paratergal lobe. The sclente Su is united with tonofibrils marked 117 passing through both layers of paratergal ectoderm to insert on the outer paratergal cuticle. These fibrils are not striated, they are confined to the ectoderm and cannot be muscular (cf. Barlet, 1967, muscle 117) and they stain as do other tonofibrils, brown with Mallory. Level D shows the origin of the pleural apodeme from the pleurite. The strengthened lateral coxal margin (not a trochantin) is now separated from the pleurite by an expanse of flexible ventral (sternal) cuticle, On the pleural apodeme insert the longus depressor muscles from the nochanter, dep.tr.~ (see also levels E and F), the intrinsic d-epressr to the trochanter insert on the proximal mesial face of the coxa, dep.tr. Muscle 64 is shown in its entirety in level E and probably corresponds functionally with the superficial pleurals of JQPYX, figure 10 and/or the superficial pleurals of Tomoceius figures 20, 24H, 25K, 27P, R. The anterior part of the insertion of the coxal retractor muscles, sectors 103 and 104, is seen dorsally, the muscles slope postero-ventrally through levels E and F to their origin on the posterior and mesial margins of the coxa in G. Level E Through the middle of the coxa, the pleural apodeme is well invested by the insertion of dep.tr.1. and dorsdaterally the small muscle 116 is probably a stabilizer. The origins of the long coxal protractor 101, and of the short coxal abductor from the lateral coxal margin to the pleural apodeme shown, and drawn at a higher magnification in E, as indicated by the rectangle. The other muscles have already been mentioned. Level F. Through the postereventral termination of the pleural apodeme, attached here to connective tissue linking with the median tendinous endoskeletal plate (marked and shown in black) and tonofibrillar connectives to the sternal cuticle. The lateral coxal margin is separated from the pleurite by an expanse of flexible membrane from which arises the coxal abductor 102; the pleural retractor 115 similarly leaves the flexible membrane above the pleurite and passes posterdorsally through level G to the tergite. A massive coxal adductor 86 arises as shown and inserts on the median endoskeletal plate. The intrinsic levator and depressor muscles to the trochanter lev.tr. and dep.tr. are shown inserting on the lateral coxal margin (left) and cut within the mesial coxal cuticle (right). The longus dep.2r.l. muscles form two main masses passing out of the coxal cavity behind the level of the pleural apodeme. Level C. Through the hinder limit of the coxal origin. The pleurite and lateral coxal margin are closer together, but there is no firm articulation between them. The other muscles shown have already been mentioned.
144 346 S. M. MANTON LEPISMA a l axis of promotor remotor swing of coxa katopleure anopleure b 9 articulation a. Figure 33. Lepisna sacchanna L. The inner surface of the mesothoracic cuticle, after Barlet, The tendinous endoskeletal plate and its connectives are marked by interrupted lines. The close unions between coxa and aochantin, articulation a below and fi above of Barlet, are marked by circles a and b and labelled as (b). The limits of the three pleurites are indicated by the black zones in (b), but in reality they overlap one another and do not form a pavement, see figure 35a. The posterior indentation on the coxa cames the tendon of an adductor muscle (figure 35e, muscle 106). The points marked by circles limit the coxal swing to one about the axis shown in (a) but a true articulation is only present at articulation b (see figure 35d).
145 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 347 LEPISMA I a axis of promotor remotor swing of coxa b Figure 34. Lepisma sacchanna L. (a) Lateral and (b) ventral views to show the positions of the legs and their movements. The three sternites slope postero-ventrally and carry posteremedian and lateral flanges, as shown. The lateral flanges lie ventral to and against the coxae. The coxal origin from the body is roughly indicated by the heavy dotted line at the anterior end of left coxa 2 in both figures. The coxae arise ventro-laterally and lie in grooves between the paratergal lobes and lateral flanges of the sternites. The coxae are much flattened, cf. other legs in figures 9, 12, 18, 30, 31, and slope postero-ventrally. Owing to the position of each leg, the small promotor-remotor movement of the coxa about the axis shown combines with the adductor-abductor movement of the trochanter on the coxa to give the main stepping movement, a condition found in no other hexapod (see further in text). The morphology of the coxa-trochanter joint is such that the trochanter and femur swing forwards dorsal to the coxa, and most extensively on leg 1, see also figure 35F. The arrowed line crossing leg 1 in (b) shows the very great angle of swing of the telopod. The points F, C, H on (b) show the levels of the sections in figure 35 relative to the coxae. The preparation of the sections has caused some shrinkage and therefore the tergites are telescoped, in contrast to their positions here in (b).,
146 348 S. M. MANTON H 2. \ skeletal 1tru1 1W.W. dcp.cr C 4 d mr. e
147 EVOLUTlON OF HEXAPOD LOCOMOTORY MECHANISMS 349 Figure 35. Lepisma saccharina L. (a-c) Diagrammatic transverse sections to show features of the mesothoracic leg base and sternite. The three levels F, G, H are marked on figure 34b and relate to the coxa and pleurites, but shrinkage of the specimen during sectioning results in some telescoping of the segments SO that the anterior margin of the metathoracic tergite is cut above the mesothoracic leg base in G; the natural positions of these structures are shown in figure 34. The vicinities of the tergites and sternites are stippled in a contrasting manner and the coxal cavity is marked by interrupted lines. The downward slope of sternite 2 is shown by the three diagrams, and the boxing in of the coxal base by the lateral flanges of the sternite and by the paratergal lobes. The anterior part of coxa 2 is cut in (a), the middle of the coxal base in (b) and caxa 2 lies free and horizontally in (c), sternite 3 now lying above it. In (c) the pleurites and pleural apodeme situated in front of leg 3 are shown, they are similar in form anterior to level F (a). The insertions of the long levator nochanteris muscles at the tip of the pleural apodeme and on the anopleure are shown in (c), but other muscles are omitted. The wide connective tissue endoskeletal sheet forms a plate above the nerve ganglia (b) and narrows to its median connection with the sternite between the nerve cords in (c). (d) Diagrammatic thick transverse section to show: above, the details of the pleural apodeme and its muscles (as on (c) but on the mesothoracic segment) and below, the coxal connections at a slightly more posterior level, near F above. The anterior part of the coxa has a point of close union with the nochantin above, and a less close one below, articulations a and b (a and p of Barlet, 1951, and marked by two rings on figure 34 here). The only true articulation is articulation b. The arrow indicates the intucking which forms the pleural apodeme, a highly sclerotized thickening of the ventral wall. The apodeme bears: muscles to the tergite above (muscles 163, 164 of Barlet, 1954), laterally directed muscles to the tergite and to connective tissue crossing the mouth of the paratergal lobe, and the long levator muscles from the trochanter leu.tr.2.k.a. (muscles 193, 194 of Barlet, 1954). Flexible, little sclerotized pleuron forms bulges (marked by interrupted direction lines), on parts of which lie the trochantin and katapleure (marked by heavy direction lines). The coxal details shown lie just posterior to level F above. The trochantin is separated from the coxa by a thin, short cuticular link at b, staining blue with Mallory s triple stain, the rest of the coxal cuticle staining red. At Q the link is more ample, and posterior to these points the trochantinal sclerotization ends. There is no third articulation of Barlet posterior to the coxal groove carrying the adductor muscle 106 and its tendon, the pleuron here bearing no sclerite. (e) Section a little more posterior than (d) passing through the coxa almost as in (b). The sclerotized coxal margin is not simple, a stout marginal zone bears an adductor muscle (1 10 of Barlet, 1954). ventral to this is a groove in the coxal cuticle giving rise to the tendons bearing adductor muscle 106 of Barlet, and below this a thud adductor muscle, 108 of Barlet, arises from the lateral face of the coxa, stout tonofibrillar connections to the cuticle being in evidence at the origins of these muscles; 106 inserts on the endoskeletal plate of the prothorax, 108 on the endoskeletal plate of the mesothorax, mainly on the ventral face near the middle line, i.e., as long a muscle as possible, and muscle 110 inserts on the sternite, near the middle line, all long muscles. The long levators from the trochanter, Zev.tr.2.h. (muscles 193, 194, Barlet, 1954, to the pleural apodeme) and leu.r*.lan. (muscles 195, Barlet, 1954) are hatched, see also (c) and (d)), separate within the coxa from the levator nochanteris group of fibres marked lev.tr. in (c) and pass upwards to insert on the katapleure and anapleure respectively in (c) and (d). The cut, hatched, muscle at the entrance to the paratergal lobe is the longest of the dorsal longitudinals (muscle 10 of Barlet, 1953).
.")
148 Figure 36. Lepisma saccharinn L. Leg morphology and movement. (a) Left leg 2 in ventral view, the overlapping sternal flange has been removed but the paratergal Lobe is in place. The leg lies in its natural position, the coxa sloping posteriorly and slightly downward from the body (see figures 34, 35). The morphologically anterior face of the coxa becomes the apparent ventral face. The proximal posterior rim of the coxa, at its union with the body, is shown by the dotted line. The intrinsic muscles of the leg are drawn, but only an indication is given of the extrinsic muscles (see Barlet, 1954). The trochanter articulates into a postero-dorsal concavity on the coxa, the parts being so shaped that the femur, when elevated, lies against the dorsal posterior face of the coxa, between the coxa and body. The heavy dotted outline and the stippled face of the telopod show the elevated position (see also figure 35a). The Ievator-depressor movement of the trochanter on the coxa swings through a very large angle, over 90". as shown by the contrasting positions on (a) and (b) and the arrowed, interrupted line. This movement of the trochanter becomes the largest component of the promotor-remotor movement of the limb-tip, and is obtained without large emarginations, such as needed for wide coxa-trochanter movements in Symphyla (Part 9, figure 6) and Chilopoda (Part 8, figures 58,63), or the extreme specializations of the few diplopods with comparable mobility (Part 6, text-fig. 12 and 13). (b) Outline of part of the left prothoracic leg in the same view as (a). The coxa is similarly backwardly directed, yet the range of movement of the limb-tip is as in Pehobiug figure 7a, which is forward of the coxa-trochanter joint. This range of movement is made possible by the head of the trochanter of leg 1 being even more curved than that of leg 2.
149 P./// C b C B / '*W j i! p. ' a - T t *4.* ",.' '-1!' d e Figure 37. Petrobius brevistylis Carpenter. Length 10 mm. Tracks left on smoked paper by the jumping escape reactions, printed in reverse. The marks left by thoracic leg 3 and by about six abdominal coxosternal plates are labelled on (d). (a) Site A is one of landing from a previous high jump and the take off for the next which lands at B. Another take off from B springs the animal into the air to land at C, from which there is no further high jump and the animal ran off by an irregular performance of the normal jumping, non-airborne, gait. Only sites A and B show marks left by the springing abdominal coxosternal plates. (be) Sites of landing and take off for high jumps on a larger scale, the length of the animal being shown by the marked 10 mm. line on (e). For further description see text.
150 352
151 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 353 Figure 38. Petrobius breuistylis Carpenter. Skeleto-musculature of the abdomen. (a, b) Diagrammatic transverse sections through abdominal segment 111, at the levels indicated by the arrows on (c). In (a) abdominal tergite 111 and coxo-sternal plate 3 complete the circumference; the dorsal anterior margin of tergite IV is also cut where it carried muscle 20, see the longitudinal section of the same segments shown above in (d). Only some muscles are drawn, see figure 39. (b) Transverse section at a more posterior level (see c, d) where the whole of the anterior end of the exoskeleton of abdominal segment IV is also cut, coxo-sternal plate 3 now lying free from the body and covering the anterior end of caxosternal plate 4. (c) Ventral view showing the nature of the overlaps of the exoskeletal structures. The supposed lateral junction between coxa and sternite, line Z, is drawn in section in (e). (d) Longitudinal median section through a partially flexed abdomen, to show the nature of the overlaps of the parts on dorsal shortening and the layout of the trunk musculature. The two components of the endoskeletal system, the coxal tendon and the lateral endoskeletal tendon complex are marked on the right, but the latter is much more complex than shown. On segment IV are marked: the dorsal muscles 14, 5, 8-11, 12-13, each group being indicated by one line, muscles 6, 7 are separately drawn, as are the ventral longitudinal muscles 14, 15. From the lateral endoskeletal tendon complex at the anterior end of segment IV are shown the deep dorso-ventral muscles 20, 21, and the deep obliques The rest of the deep obliques are shown diverging from the following lateral tendon complex: muscles 23, 24, 25A, 25B, 26. Muscle 2SA, 2SB curl round muscles 24, 26 as indicated. All these muscles are segmentally repeated, but only muscle 26 is drawn twice. The anterior and posterior proximal margins of the coxo-sternal plate of segment 111 are marked E, F respectively, as on (c). The heavy arrow within segment VI indicated a probable path of hydrostatic pressure at a jump exerted against the ventral face of the coxo-sternal plate, posterior to its muscles from the body. (e) Section through the junction of sternite and coxa, line 2 in (c), to show the fold in the cuticle. Scales are cut external to the cuticle and ectoderm internal to it.

152 3 54 S. M. MANTON
153 Figure 39. Perrobius brevistylis Carpenter. Sagittal view of a longitudinal half of abdominal segments IV-VI to show the abdominal musculature, redrawn from data kindly provided by Dr G. E. E. Scudder. Cut tergites and the stiff sclerotized cuticle of the coxo-sternal plates are shown by heavy black lines. The overlapping coxo-sternal plates 3-5 from segments 111-V are shown below, the numbers being situated on the styles. The lateral endoskeletal tendon complex is indicated diagrammatically and labelled on the right, that crossing intersegment V-VI is cut short to expose the more external muscles. The coxal tendon, situated in the segment in front of the main part of the lateral tendinous complex, is shown segmentally. The dorsal tendon anchoring the dorsal muscles to the cuticle is shown on segment V. The dorsal and ventral longitudinal muscles and the deep dorso-ventrals and deep oblique 26 are shown on segment IV, as on figure 38d, other deep muscles being omitted. On segment V the deep dorso-ventrals 20, 21, and deep oblique 26 and ventral longitudinal muscle 14, attached to rhe tendons crossing intersegment IV-V, are partly cut away to expose the rest of the deep oblique muscles: 23-24, 25A, 258, 26, muscles 25A. 25B twisting round muscles 24, 26 as shown diagrammatically in figure 38d. Only part of muscle 23 from this segmental tendon is shown, but the entire muscle 23 from the following tendon is drawn crossing most of segments IV-V. Muscle 23 ends on the tergite at the termination of muscle 25B. On segment VI are shown the dorsal oblique muscles 6, 7 and muscles 27-29, from the lateral part of the coxo-sternal plate anterior to the coxal tendon, which insert on the lateral part of the tergite. Muscles 15, 20, 22 drawn on segment IV represent the normal segmental complement of deep dorso-ventral, deep oblique and ventral longitudinal muscles. The additional mucles in these categories, 21, 23, 24, 25A, 25B, 26 mainly shown on segment V but terminating from segment I11 to intersegmental tendon VI-VII, are concerned with the abdominal jumping mechanism, see text. w VI VI

154 3 56 S. M. MANTON 10. GENERAL CONCLUSIONS BASED ON THE ANALYSIS OF HEXAPOD MOVEMENTS, HABITS AND ASSOCIATED MORPHOLOGY AND THE PROBABLE EVOLUTION OF THESE CLASSES A review has been given of the many ways in which hexapodous arthropods use their legs, together with the body structure which makes these movements possible. It remains to show the bearing of this data upon our knowledge of arthropodan evolution. Direct fossil evidence of the origin and evolution of hexapods is far too scanty to give a precise picture. It is limited to the earliest known collembolan, Rhyniella pruecursor Hirst & Maulik, Devonian, an already specialized member of its class, Scourfield, (1940) and Jeanne1 in Grasse, (19491, and the Monura, Lower Permian, with close thysanuran resemblances (Sharov, 1957). There are also many examples of fossilized Pterygota from various orders, all too advanced along their own lines to give any information concerning ancestry of classes. 10A. THE EVOLUTION OF THE HEXAPOD THORAX The thorax of the hexapod classes consists of three thoracic segments, usually bearing the three pairs of thoracic legs. Thoracic segments, differentiated from the rest, are present only in association with well developed legs.* When these legs are very short or absent, as in many pterygote larvae, the thoracic segments are not so well differentiated from those following. Where legs are long and strong, in either larvae or adults, they need: strong insertions on the body supporting whatever type of coxa-body articulation may be present, and an abundance of suitably supported extrinsic muscles to work the legs. Where these requirements are absent there is no well differentiated thorax, see Part 8, pl. 5, figures , showing the well-sclerotized limbless thoracic segments of the larvae of Anisopus (Diptera) forming an even series with those behind them. As noted in Part 2, the advantages of a hexapodous state in insects, crustaceans and arachnids is a fanning out of the fields of movement of the legs, so that there is little or no overlap (figures 5j, 6m, 7a). The animals can then use a wide range in gaits, an assistance in changing speed (Part 2, Table 1). When fields of movement scarcely overlap, leg length can be greater without incurring mechanical interference between successive legs (cf. Scutigera and Ligia, Part 2, text-figs. 2, 3). Longer legs can increase the stride length, provided the angle of swing of the leg remains the same, and if the frequency of stepping also remains the same, the speed of running is augmented (8 4). It is fundamentally necessary for a small number of long legs to arise close together. If they were spaced far apart on the body, the legs of a pair, usually used in opposite phase, would throw the body into undesirable undulations which would have to be controlled for efficient locomotion. Legs arising close together just behind the head also serves to support the mouth parts and feeding movements. Abdominal limbs, posterior to the thoracic legs, have persisted in so far as use has been made of them. They act as stabilizers, assisting thoracic running in It has been maintained by Tuxen (1970) that the distinguishing character of the Hexapoda is not that they have six legs but that they have a thorax of three well defied segments. This suggestion takes no account of function.
155 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS Petrobius, Campodea and possibly Lepisma, their tips touching the ground. Abdominal limbs are associated with jumping in many ways, the coxo-sternal plates of the Machilidae providing the points of take off for the abdominal jumping mechanism; the springing organ and the hamula of Collembola are believed to be modified limbs. And there are the better known gonopods; etc. Thoracic segments are often fused together, giving great stability to the leg bases of Pterygota. Here also is maximum sclerotization of the thoracic exoand endoskeleton, so providing a strong base for flying movements. But in the jumping Collembola the pro- and mesothoracic tergites are fused (8 9A(vi)).* The increased rigidity so gained assists the control of hydrostatic pressure at the jumps, and has permitted a simplification of the dorsal longitudinal muscles of these two segments, so making space for extrinsic leg muscles. In particular, those of leg 1 insert on either side of the now rigid intertergal joint (figure 18a). But mobile inter- and intrathoracic jointing is not incompatible with the advantages of a thorax. Such mobility is used in crevice penetration by Diplura, particularly in the Japygidae (8 6A(i)), and by Protura, and the mobility is less in the Campodeidae where the running ability is striking ( 8 6A( iii)). A flexible thorax is associated with: the presence of well sclerotized tergites and sternites, a wide expanse of flexible pleuron between them, many pleurites, and laterally directed coxae articulating at least with the sternites. This flexibility and associated morphology exists in the epimorphic Chilopoda as well as in the Diplura and Protura, and probably has functional rather than phylogenetic significance. The converse morphology exists in many Diplopoda and in the Collembola and Thysanura. Here a tergite or tergo-pleurite envelops the trunk or the thoracic segment, the pleuron is small and not freely exposed, the pleurites are few and the sternite absent in the two hexapod classes. These contrasting features are associated with a need for rigidity. Since the morphology, conferring either mobility or rigidity, appears to be basic for each class, it is probable that neither the one nor the other should be considered primitive, both being alternative types of evolutionary advance. There is more resemblance between meso- and metathoracic segments than between either of them and the prothorax. Tuxen (1970) notes that there are no wings in the Apterygota to account for this. However there are plenty of reasons for the differentiation. (i) Where there is no narrow neck behind the head, as in caterpillars, the prothorax may be shorter than the following segments, but otherwise is generally similar, and in the limbless Anisopus larva (see above) the prothorax is very much like the meso- and metathorax. But where a mobile narrow neck is present behind the head, the prothorax must either taper anteriorly, as in most apterygotes (pls. 1-6), or possess projecting shoulders. (ii) The prothorax is smaller than the meso- and metathorax because it needs to house a smaller bulk of muscles. The prothoracic legs are shorter and have different fields of movement from those following, and they need a smaller bulk of extrinsic muscle, a little differently arranged from those on the meso- and metathorax. The thoracic tendinous endoskeleton is essentially intersegmental in origin and bears the dorso-ventral and deep oblique muscles which insert on the tergitc behind. There are no comparable tendinous The statement by Tuxen (1970) that in the Collembola the prothorax vanishes dorsally is incorrect, see 5 9A(vi). 357
156 358 S. M. MANTON processes passing posteriorly from the neck region and there are no prothoracic deep dorso-ventral and deep oblique muscles (figures 13b, 21a). These and other reasons account for the differences between the prothorax and the mesoand metathorax in the apterygote classes. Thus a functional consideration of thoracic structure indicates that the differentiation and enlargement of three pairs of legs must have initiated the evolution of the thoracic segments in all hexapods, and not vice versa, as implied by Tuxen (1970). 10B. THE EVOLUTION OF HEXAPOD GAITS AND SPEEDY MOVEMENT Quick running by the use of fast patterns of gait, as in the fleeter myriapods, does not occur in hexapods. Stability requirements generally necessitate that there be no unstable moments in time when the body of a hexapod is supported by only two limbs (0 4A). Usually only the slower patterns of gait are practicable to a hexapod, with a backstroke of no shorter duration than the forward stroke. A certain measure of similarity exists between the use of the legs of all hexapods for this reason. But the gaits employed by the several hexapod classes are not as uniform as implied by a consideration of the Pterygota alone (Hughes, 1952). The Diplura Campodeidae are characterized by great agility and length of stride, momentarily unstable gaits being employed at their fastest running. The Japygidae, on the contrary, are not good at surface locomotion, but excel in the negotiation of tortuous crevices by strong leg movements. The exact use to which the proficiencies of the Campodeidae are put is probably indicated by their being the last to leave a tullgren funnel, yet they are the first to desiccate outside it. The agility here is used to find, keep or change the surrounding microclimate in soil spaces (J. G. Blower, pers. comm.) The Collembola avoid using momentarily unstable gaits when running and achieve speedy locomotion by jumping with the abdominal springing organ and not by running. The springless deeper living Collembola appear to have been derived from jumping ancestors, their springing organs and associated morphology being secondarily altered (0 9B). The whole trunk and limb morphology of Collembola appears to be correlated with the jumping mechanism, worked by hydrostatic propulsion, the pressure being generated momentarily by the trunk musculature. The Thysanura show some of the most specialized leg movements existing among hexapods. Fast progression is achieved in the Machilidae by jumping gaits which are alternative to the more normal gaits used on the underside of rocks, etc., and in the Lepismatidae by a unique leg action (0 7C, C(iii) and below) based upon that of the Machilidae. It has been shown in previous Parts how varied and successful are the myriapodan methods of speedy running, used when venturing over exposed surfaces. The hexapods, too, have needed speed when they have not escaped by going deeper into the soil. The Pterygota gain speed by flight, but on the whole their plantigrade walking is more leisurely than in the apterygote classes. Speed is gained in the Diplura by the campodeid specializations, in the Collembola by jumping escape reactions, and in the Thysanura Machilidae by normal hexapod walking gaits, by highly specialized jumping gaits and by high jumping escape
157 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 3 59 reactions. The very rapid runs of short duration in the Lepismatidae are based on a probable derivative of the machilid jumping leg movement. All these feats depend upon facilitating morphology, differing from class to class. A need for speedy running is avoided by the habits of Japygidae, the non-jumping Collembola and Protura in association with living under cover, and there are many comparable examples among both adult and larval Pterygota. Speed of running is affected by leg length, by the angle of swing of the leg, by the relative duration of the backstroke and by pace duration (see above). Fleetness results from rapid stepping in many of the smaller Pterygota, and occurs in extreme form in the Lepismatidae, where leg morphology leads to this end. A large angular swing of the leg in Lepisma is restricted to the telopod which is slender and lightly constructed. The coxa moves far less, but provides housing for the bulk of muscles needed to work the distal wide and rapid swing of the leg. In no other arthropod has a limb movement been found in which the promotor-remotor and the adductor-abductor movements are almost merged and the angle of swing to differ along the leg as in Lepisma (8 7C(iii)). The legs of the Lepismatidae are as far removed from a primitive type of hexapod leg as are the hind legs of jumping mammals from the primitive mammalian type. Yet the Lepisma leg-base has been hailed as primitive in entomological literature, without any consideration of function (Denis, 1949; Matsuda, 1960, 1963, etc. and see below). The patterns of gait used by extant arthropods which could have given rise to the gaits of hexapods are few. It has been shown in Part 9 that the symphylan gaits represent the slower end of the series seen in the epimorphic Chilopoda. Both ends of this long series (Part 3, text-fig. 3 right and Part 9, figure 1 left) require structural specializations for their execution. The middle gaits, representing the faster of the symphylan gaits and slower of the epimorphic chilopodan gaits (Part 3, text-fig. 3 left and Part 9, figure 1 right), are less differentiated, resembling the slower gaits of the Pauropoda (Part 3, text-fig. 12a), and the faster gaits of many hexapods figure 3. Most of the myriapodan gaits, with the associated morphology, are much too advanced along their own lines to have had anything to do with ancestral states leading to hexapod gaits ; the facilitating morphology of the conspicuous myriapodan locomotory achievements does not lead towards the morphology of hexapods. Even the Symphyla, in employing their slow patterns of gait, are highly advanced in their remarkably rapid forward stroke of the leg and associated morphology, which is the basis of their agility and stability in running, in spite of great flexibility used for other purposes (Part 9). The middle-gear gait of the Onychophora (Part 1, text-fig. 6) resembles the simplest of the myriapod and hexapod gaits in most respects, but the blunt limb tip of the Onychophora necessitates leg n + 1 being put on the ground just after leg n is raised. With the advantage of a narrow sclerotized tarsus and a unguligrade stance, the footfall of leg n + I can take place simultaneously with the raising of leg n or even before this event, and on almost the same footprint, so that no moment in time exists when the body is unsupported by one or other of two consecutive legs, a very important matter to a hexapod (cf. Part 1, text-fig. 6b, with Part 3, text-fig. 3 and figures 2, 3 here with long time intervals k, and figures 6a here where the change in footfalls is simultaneous). Therefore it may be suggested that hexapod gaits arose in animals with

158 360 S. M. MANTON increasing length of legs and pointed narrow limb tips, but lacking the advances seen in present day myriapods; and that the evolving hexapod gaits could have originated from those seen today in the onychophoran middle gear with the additional advance of improved stepping following on the sclerotization of the leg. But here the similarity between the gaits of hexapods ends. In each of the five classes the legs are used in different ways, which, as shown above, stand out from the common features resulting from using only three pairs of legs. These differences are such that it must be supposed that the hexapodous stepping advanced independently in each class, along with the facilitating morphology (see also 0 10E, G). 1OC. THE EVOLUTION OF EXTERNAL THORACIC STRUCTURE OF THE HEXAPOD CLASSES The basic requirements of a thorax are the provision of: a suitable insertion for the leg; adequate support for the coxa-body union and for extrinsic muscles controlling leg movements; and flexibility or rigidity as demanded by the needs and habits of the animals. These requirements are met differently in the several hexapod classes. (i) Thoracic rigidity and flexibility The Diplura show conspicuous flexibility of thorax, both inter- and intrasegmental, and the Protura are flexible also (see above, 8 1OA). An expanse of flexible pleuron carries many pleurites (8 6A(i), 7A, A(ii), D). Greater thoracic rigidity is gained in other hexapod classes by: the presence of shorter, wider segments; no narrow inter-segmental junctions; by enveloping paratergal extensions from the tergites (Thysanura and Collembola) ; and sometimes by tergite fusion (on pro- and mesothorax in jumping Collembola). It is possible that collembolan tergites represent undivided tergo-pleurites. The pleurites are few where there is IittIe thoracic flexibility, one to three in Thysanura, one on the metathorax only in jumping Collembola, and the functions served by these pleurites are various (0 7). (ii) The insertion of the legs The leg-base is remarkably different in the hexapod classes. A sternal articulation with the ventral coxal rim is present in the Diplura, but without the rocking movement so essential to most Myriapoda; a corresponding rocking takes place within the leg itself. The proturan coxa possesses articulations both with the sternite and a pleurite, together with a coxa-sternite rocking mechanism, quite differently contrived from that of myriapods. Only one, pleural, articulation with the dorsal rim of the coxa is present in the Thysanura, but unlike the Pterygota, the pleurite providing the articulation is mobile, hinged to the tergite in the Machilidae and supported therefrom in the Lepismatidae, in contrast to the fixed pleurite forming the principal articulation with the pterygote coxa (Part 9). In the Collembola there is no articulation between coxa and body because such a structure would be mechanically unsuitable. Instead there is an elaborate suspensory system of the leg from the endoskeleton, which resists dislocation at the joints, or any
159 EVOLUTION OF HEXAFQD LOCOMOTORY MECHANISMS 361 adverse effects of momentarily increased internal hydrostatic pressure at a jump (figure 30). These four entirely different types of leg insertion are not derivable one from another, but represent the meeting of hexapod needs in different ways and must have evolved independently. (iii) Pleurites, subcoxal segments and fright The composition and evolution of the pterygote pleuron and the supposed evidence for and against the view that subcoxal segments, one or two, have been flattened and incorporated into the pleuron, or vice versa, has been discussed many times; Verhoeff (1902), Borner (1921), Crampton (1926), Snodgrass ( 1927), Hansen ( 193 O), Ferris ( 1940), Carpentier (1946), Denis (1949), Weber (1928, 1952), Gouin (1959), Matsuda (1960, 1963) and others. Attempts have been made to reconcile the conflicting views, e.g. Gouin (1959). Snodgrass (1927) admits that proof of his theoretical derivation of the insect (pterygote) pleuron is lacking. Weber and Matsuda show how the various pterygote pleura might have been derived from two concentric sclerotizations around the leg base, named the katapleure and anapleure. Matsuda (1963) notes that an initial subcoxa might have existed as a simple ring surrounding the base of the leg, and that later it may have broken up into sclerotized areas, the pleurites, forming two rings above the coxa as seen in some Apterygota. But two pleurites above the coxa are present only along the dipluran thorax where there is a diametrically opposite type of coxa-body junction to that of the Pterygota; on the prothorax only of the Thysanura Machilidae but not the Lepismatidae, and again there is no functional similarity between the leg bases of Thysanura and Pterygota (figures 9b, 3 lb, 32A); and in the Protura where there are dorsal and ventral coxal articulations, arranged as in no other hexapods, and where the two pleurites provide a unique coxal rocking mechanism. Matsuda s view that some general desclerotization in hexapods might account for the breaking up of originally highly sclerotized areas has little in its favour as an explanation of apterygote pleurites. Large animals, of mechanical necessity, are relatively more highly sclerotized than small ones. The five foot terrestrial Arthropleuru arrnuta, late Carboniferous (Rolfe, 1966), is much more sclerotized than small present day terrestrial arthropods. Heavily armoured fossil fish gave place to lightly armoured ones of the same size, but there is no evidence that terrestrial arthropods were similar in this, or that their sclerotizations, relative to size, were greater than they are now. But degrees of sclerotization, or of stiffening of the cuticle, differ greatly in respect of habits, e.g. burrowing Diplopoda in contrast with other myriapods. The comparative maps of the pleuron and its supposed derivation (Weber, Matsuda, etc.) are valid only for the pterygote orders. Here the subcoxa, if there ever was one, becomes synonymous with the pleuron, which enlarges, leading to the vanishing point externally for the sternite, so giving the strongly sclerotized lateral thoracic support for the wing base. But the origin of this pleural sclerotization is another matter, and the myriapods and apterygotes do not provide the evidence claimed of them by entomologists, mainly because their leg-bases and pleura have been inadequately described hitherto and no functional concepts have been considered. Therefore there have been no known
160 362 S. M. MANTON reasons why hypothetical subcoxal rings of sclerotization round the leg of a myriapod or apterygote or primitive insect should not have been a working proposition. But the functional information now available on these animals renders the above concept untenable because it constitutes a mechanically unsuitable leg-base. An animal must be a working whole at all stages in its evolution. Where foundations are at fault, edifices of theory and belief must fall. Some of the foundations which are here considered to be invalid, are as follows. The view that arthropods are monophyletic, and that the insects and crustaceans are closely or distantly related, is now untenable. The fundamental differences in the evolution of the jaws of Crustacea and of the Onychophora-Myriapoda-Hexapoda assemblage indicates an independant evolution of these two groups (Manton, 1964), a conclusion amply supported by embryonic fate maps, etc. (Anderson, 1966, 1969, in press) and by other evidence. Thus resemblances and differences between the coxae of Anaspides or of an isopod and of insects becomes totally irrelevant, unless convergent features are looked for. The view that the five hexapod classes constitute orders of a single taxon, the insects, in tenaceously held by many, although questioned by others, and results in the claiming of homologous structures throughout the five groups, such as pleurites, subcoxal segments, endoskeletal units (see below) etc. These interpretations are not upheld here. The existing naming of pleurites in myriapod and hexapod classes does not indicate homology. The so-called trochantin of Lepisma, which carries the only articulation between coxa and body, can hardly be homologous with the pterygote trochantin in which class the principal coxal articulation lies with the fixed pleurite, not with the trochantin. The myriapodan katopleure lies closest to the coxa, as does the trochantin in apterygotes, but the myriapodan katopleure never bears a coxal articulation and cannot be homologous with the apterygote katopleure lying further from the leg than the trochantin, etc. It is concluded here that pleural sclerotizations are formed in apterygote hexapods, as in myriapods, just where they are required for mechanical reasons and that there is no common plan, because leg insertions on the body differ profoundly from class to class. The view that the nature of pleural morphology can be ascertained by careful scrutiny, or by mapping the pleuron into areas bearing the names of sclerites is here considered to be fallacious. The limits of pleural sclerotizations, and the presence or absence of articulations, can be ascertained with certainty only by reference to serial sections stained with Azan or Mallory's triple stain, which show exactly where the different degrees of sclerotization lie. Thickness of cuticle is no indication of the amount of sclerotization. Cuticles stained with chlorazol black may also be of great service. There can be nothing fundamental about the three claimed coxa-body articulations in Lepisma when in fact only one exists. How the three could function, when their claimed positions are not in line with one another, has never been considered, Arguments fail which are based upon these three supposed articulations. Similarly the identification as an anopleure (Barlet, 1967) of a functionally essential short link of flexible pleuron between the tergite of the Machilidae and their single meso- and metathoracic pleurites (figures 31, 32F, G) is invalid. This distortion of the facts attempts to force a homology with the Lepismatidae, but even if valid,

161 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 363 this claim does not prove that the Machilidae have 3 pleurites as have the Lepismatidae. The view that the pleurites of Lepisma, however graphically attractive they may be, illustrate a basic or primitive condition, which may have led to that of the Pterygota, is unsound. These pleurites form a vertical imbricating series anterior to the coxal rim and support coxal and telopod movements which are unique among known hexapods. The changing degree of overlap of the pleurites supports the coxa anteriorly during its rapid swing almost in a horizontal plane, the posterior coxal rim deforming the pleuron. The coxal promotor muscles, which have shifted in part to the pleurites of Petrobius, originate entirely on the pleurites of Lepisma, where they serve the same purpose of coxal promotion. Such pleurites and coxal movements are not present in any other class, and pterygote coxae do not lack promotor muscles. It is concluded that hexapod pleurites do not correspond from class to class, either in morphology or in functions. Neither do they correspond with the various arrays of myriapodan pleurites. Moreover in the Diplura, Collembola, Protura and Thysanura the thoracic pleurites differ from segment to segment on the same animal, as in many myriapods. The pleurites of the Thysanura, although showing the nearest approach to those of the Pterygota in their articulation with the dorsal and not the ventral rim of the coxa, are committed to most important mobility in respect of functional needs, while rigidity of the main pleurite is required by the Pterygota. These opposite conditions of the coxa-pleural joint can only mean evolutionary divergence of Thysanura and Pterygota. The inexact superficial resemblances between Protura, Diplura and Chilopoda, all possessing more than one pleurite, but only the proturan trochantin articulating with the coxa, is related to trunk flexibility served by a body form possessing a large sternite and a coxa-sternite articulation, a shape suited by rocking leg mechanisms. The rocking is differently contrived in Myriapoda, Protura and Diplura. The general form of the segments in these hexapods is not considered to be primitive (see above 5 loa), and inexact pleurite resemblances cannot constitute a basic or primitive condition. Lastly, the arc-shape of proturan pleurites is related to their movements, there is no good evidence that this shape indicates that the pleurites are the remains of subcoxal segments. The view that subcoxal segments can be seen in extant apterygotes is not upheld. The Collembola do not show either a simple, or a primitive, leg-base as has been claimed (Denis, 1949). The pleural bulges above leg 1 are not subcoxae, but outbillowings of flexible cuticle between the endoskeletal ties which hold the pleuron under control at the jump when internal pressure is momentarily increased. The partial division of coxa 3 and the complete division of coxa 2 has nothing to do with subcoxae or pleuron, but is directly concerned with functional needs dependent upon the jumping mechanism and leg suspension. The present work is in disagreement with the identification in Collembola of anapleure, katapleure and trochantin given by Carpentier (1949). The latter is shown to be part of the true coxa on the meso- and metathoracic legs and not to be a sclerite at all over leg 1, while the claimed anopleure is an area of undifferentiated pleuron ($ 7B(ii), 9A). There is only one true pleurite on the thorax of Tomocerus. It is situated above leg 3, where it supports the posteriorly directed field of movement of this long leg.
162 364 S. M. MANTON Thus no myriapod or apterygote class provides any evidence supporting the supposed origin of the pterygote pleural sclerotizations from concentric sclerites round the leg base in any non-pterygote classes. But the very real differences between the massive pleural sclerotizations of the Pterygota and all other hexapod classes indicates an evolutionary divergence of the Pterygota from the rest, which led to the evolution of wings, this needing a highly sclerotized lateral thorax. The ability to fly must have been evolved in the Pterygota from early flightless members of this class possessing considerable pleural ridigity, Tiegs (1 955) has shown how the simplest pterygote flight muscles could have arisen from coxal extrinsic muscles to the tergite, depressing the tergal walls and forming levators, the antagonistic wing depression being provided by the pleural muscles. The use of dorsal longitudinal muscles as wing depressors would follow on the formation of skeletal phragmata, so improving the simple initial wing mechanism (see also Snodgrass, 1929). Much greater advances were to follow with the evolution of new muscles; and the very rapid wing beat in some orders, combined with a minimum of thoracic deformation, is implemented by quite novel types of muscles (Tiegs, 1955). The employment of the primitive pterygote coxa-tergal and pleural muscles for flight was dependent upon the rigidity of the pleural structures, including the pleural facet of articulation with the coxa. A large pleural apodeme united with the sternal furca, the pleurite being firm above (Part 9, figure 10a), provides the necessary rigid base for flight muscles. An apterygote with a coxa-sternite articulation and no pleural rigidity (Diplura, Protura), a basically tendinous endoskeletal coxa-body union and one free pleurite on the metathorax only (Collembola), and a limb mechanism dependent upon mobility of pleurites (Thysanura, Myriapoda), all preclude the evolution of flight. 10D. THE EVOLUTION OF INTERNAL THORACIC STRUCTURE OF THE APTERYGOTE CLASSES The endoskeleton and associated muscles of the dipluran and proturan thorax have been described in 8 8 and that of the whole trunk of a collembolan has been considered in 0 9(iv), (v) and in Appendices I and I1 because of the correlations with the jumping mechanism. Detailed accounts have been given of the musculature and endoskeleton of the thorax of various thysanurans and Collembola by Barlet, 1946, 1951, 1953, 1954, 1967, Carpentier, 1946, 1949 and by Carpentier & Barlet, (i) Endoskeleton of the thorax The endoskeletons of the Diplura, Collembola and Thysanura are unlike. It has been shown above that in the Diplura the thoracic endoskeleton is largely apodemal, that it arises from sternal intuckings in both Campodeidae and Japygidae, and that a spina projecting internally and backwards from the posterior margin of the sternite, is present only in japygids, the apodemes being differently arranged in campodeids. In both groups there are some endoskeletal tendons, the details differing (8 8A). In the Protura apodemal ridges are present, differing in detail from those of japygids and lacking a hollow spina.
163 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 365 The ridges are very simple, suiting extrinsic leg musculature spread on the sternite, as in Chilopoda, but needing stronger sites for insertion, in keeping with the strong, short legs. In the Collembola the thoracic endoskeleton is entirely tendinous and much more elaborate than the tendons of the Diplura. There are no apodemes ( 5 9A(v)). The thoracic endoskeleton of a lepismatid, Lepismu saccharinu, and of a machilid, Lepismuchilis y-signatu, can most easily be compared by reference to the drawings of Barlet, 1951, Fig. 1 and 1967, Figs In both the ventro-lateral tendinous endoskeleton forms transverse bars, roughly at the intersegmental levels, bearing forward projections over the coxal cavities in Lepismachilis, and some backward projections as well. The system is linked with the cuticle by anchoring struts, which are less elaborate than those of Collembola (Appendix 11). The apodemal endoskeleton is restricted to the large pleural apodemes. There are no large sternal apodemes, and in Petrobius a prothoracic coxal apodeme replaces the pleural apodeme on this segment (figures 32A, D, E). The tendon system of arthropods is intersegmental in embryological origin (Manton, 1934) and often the serial repetition along the body persists into the adult stage, although sometimes much modified. The basic intersegmental position of the tendons is very clear in many apterygote classes and is seen, little changed, in the drawing of Lepismachilis by Barlet (1967, Fig. 1). Elaborations in shape and in secondary cuticular connections are enormously varied in the apterygotes. Apodemes, on the contrary, are not segmental in distribution. They arise anywhere, in positions where they are needed for mechanical reasons. A horizontal plate, usually above the nerve cord and below the gut, is a common feature of widely unrelated arthropods, such as Limulus (Manton, 1964, Figs ), various Crustacea (Manton, 1934, text-figs. 14, 15, 17), Diplura (figures 12, 13a), Thysanura (figure 32f and Barlet, 1967, Figs. 1-3), and the plate is below the nerve cord on the prothorax of Collembola (figure 22A). Whether the endoskeletal plate, or endosternite, is formed of connective tissue fibres, as in all of the above except the Diplura, or whether the plate is apodemal as in the Diplura and in many Crustacea (e.g. mysids, Manton, 1928, Figure 22d e.pl. ), the functional significance is the same in the provision of sites for muscle insertions. Such endoskeletal structures in roughly the same situation but contrasting in composition, can only signify the meeting of common needs by different means. And the means are so different as to imply independent evolution. An effort has been made by entomologists, e.g. Denis (1949), to find an apodemal spina or its equivalent, in all apterygotes, although it is present only in japygid Diplura (5 8A(i)). The transverse segmental tendons of the thysanuran thorax are each linked with the posterior median margin of the preceding sternite (Barlet, 1967, Fig. l), a useful tie no doubt, but not at all resembling the spina and its connections in the Japygidae. Tiegs (1955, text-fig. 1) describes muscles of Lepisma attached to what he calls a spina, but his drawing shows their origin from a shallow intersegmental junction where they are doubtless attached to Barlet s tendon system. There is no strong, hollow apodeme, based upon a thoracic infolding whose strength withstands the opposing tensions of the coxal articulation anteriorly and the coxal
164 366 S. M. MANTON retractor, and other muscles, posteriorly (figure 10). To attempt to divorce the spina from its functionally essential basal arms is comparable with expecting a head to function normally when severed from the trunk. The mid-ventral cuticular gutter along the collembolan thorax (figures 22B, 23, 24, 251, 28), is presumably the line referred to as the homologue of a spina by Gouin (1959), but there is no resemblance whatever between the two structures. The large pleural apodeme of the Thysanura has been ignored in entomological considerations of the sternal spina. Yet the pleural apodeme carries insertions of extrinsic leg muscles, as does the spina of japygids (figures 10, 32). These apodemes in the two classes are morphologically distinct, and if a spina was present in a thysanuran it would be of little use for limb musculature, owing to the position of the coxal articulation. The presence of a spina in the Japygidae, alone among hexapods, is correlated with the provision of a suitable site for coxal remotor muscles of extraordinary size and bulk, which are responsible to the very strong leg movements of these animals, used, not in surface walking, but in negotiating soil crevices. There is no apodemal furca projecting from the sternite of any apterygote class which is homologous with those of the Pterygota. There are tendinous structures which sometimes support muscles in a similar position, such as that shown by Barlet (1967, Fig. l), where a median projection from the thoracic 1-2 intersegmental tendon system extends posteriorly into the mesothorax, a clear case of parallel evolution. Thus it may be concluded that there is no basic similarity between the thoracic endoskeletal systems of the four apterygote classes, any more than between their pleural armature or their coxa-body junctions. The resemblances between endosternite positions, anchoring struts and connectives are not exact, and their general similarities are due to the mode of origin of all tendinous endoskeletal systems and to similar functional requirements. The latter are met by apodemes, or by tendons, or by both in the several classes. (ii) Extrinsic limb muscles of the thorax In all hexapods the extrinsic limb muscles form insertions upon the tergites, as in myriapods (other than Diplopoda, where it is mechanically impossible to use the tergal surface for this purpose). Some hexapod muscles insert upon the sternites, but fewer than in myriapods, but there are no sternites on the thoracic sterna of Collembola and Thysanura. Since the nature of the endoskeletal structures in Diplura, Collembola and Thysanura are so different, one from another, the insertions of extrinsic muscles on these endoskeletons, of necessity, differ from class to class. In the Diplura, bulky coxal protractor, retractor, levator and depressor muscles insert upon the apodemal system, additional to the sectors inserting on the tergites. The length of the japygid spina, and consequently the length and size of the coxal retractor ret.co.6., gives strength to the leg movements (figures 10, 11). In the Collembola most of the coxal protractors, retractors, and abductor muscles have tergal insertions, the endoskeletal system carrying the unique suspensory system of the leg, a few coxal protractor and retractor muscles and the coxal adductors (figures 22-25, 30). In the Thysanura the
165 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 367 coxal protractor, retractor and abductor muscles mainly insert on the tergite while some adductor muscles insert on the endoskeletal plate (figure 32). In the Protura the coxal protractor and retractor muscles insert on the tergal face and sternal apodematous ridges. Extrinsic muscles from the leg, arising distal to the coxa in the Diplura, are few and insert on the apodemal endoskeleton (figure 12F dep. tr.2.). In Protura they insert on the sternite. In the Collembola they are bulky and insert on the tendinous endoskeleton (figures 18, 22, 23, dep. tr. 1. ) and in the Thysanura on to the coxal apodeme (figure 32D, E, F, deptr. 1.). All are sites differing morphologically from one another. Thus there is no common basic plan in the arrangement of the extrinsic limb muscles of apterygote hexapods, their insertions depending upon the place of origin of the coxa on the body, whether the coxa is laterally directed, as in Diplura and Protura, or ventrally directed as in Collembola and Thysanura, and upon the available endo- and exoskeletal structures. (iii) Thoracic trunk musculature This system cannot be divorced from that of the abdomen, because of the functional attributes of the whole in some classes, and the manner in which the system reflects the habits of the animals. In Part 8, yp. 272, it was possible to suggest particular functions for certain components of the trunk musculature. The main components in the apterygote hexapods resemble those of myriapods, and the present analysis of structure and function has enabled further functions to be assigned to certain categories of trunk muscles. The dorsal longitudinal system connects mainly with the tergites and the dorsal segmental tendons, which may be exactly intersegmental (figures 18a, 26M) or may sag back a little distance from the anterior tergal margin, as in Petrobius. Or, when intercalary tergites are well differentiated (Japygidae), the anterior margins of both principal and intercalary tergites bear tendons of insertion for dorsal longitudinal muscles (figure 10). The dorsal systems are most elaborate where the ability to flex the thorax is greatest, as in japygids; and the formation of long muscles, crossing more than one intersegment, as in Collembola (figure 19), is associated with maintaining rigidity, as in fast running Chilopoda. A lateral longitudinal system of muscles, completing a cylinder with the dorsal and sternal longitudinal muscles in the Collembola, alone among hexapods, is most remarkable. Such lateral muscles occur in the Onychophora and in similar form in Geophilomorpha. They appear to be concerned with the sudden generation and control of internal hydrostatic pressure, in the Geophilomorpha accompanying the burrowing thrust on the soil and in Collembola for the jumping mechanism. The hydrostatic forces in both classes art' generated by pressure exerted by the trunk musculature on the haemolymph. The collembolan lateral longitudinal muscles presumably have descended from those of soft bodied ancestors, and have persisted because of the use of hydrostatic forces in the evolving jumping mechanism (see below and 8 10F). The sternal longitudinal system in myriapods, Diplura, Protura and Thysanura mainly consists of short sectors crossing one intersegment. The
166 368 S. M. MANTON formation of elaborate long sectors in the Collembola (figure 21b) is a specialization causing body shortening in association with the jumping mechanism. The superficial pleural and dorso-ventral muscles are well formed where lateral flexibility of the thorax is great, as in japygids and to a lesser extent in campodeids. Where there is no intersegmental thoracic flexibility, as in Collembola and Thysanura, such muscles are few, or absent, or diverted to the purpose shown by Collembola. Here the much enlarged pleural muscles prevent a billowing out of the body wall under increase of internal hydrostatic pressure (figures 20a, 24-27). The deep dorso-ventral and deep oblique muscles show variations in size and complexity. They appear to form two antagonistic series, the deep obliques causing slight body shortening while contraction of the dorso-ventrals causes extension. The two series of muscles together make for stability and inflexibility, as in Chilopoda, in contrast to the superficial series which promote flexibility. In Part 9, p. 119 it was noted that the Pauropoda, with rigid bodies, possess only the deep series and the Symphyla, with very flexible bodies, possess only the superficial series. In the apterygote hexapods the deep muscles are best developed in the Collembola and Thysanura, and they become particularly large and elaborate in the abdomen in association with the jumping mechanism of both classes (3 9A(iv), C(iv)). Thus the internal thoracic structure of apterygote classes is as various as are the external features, conditions which indicate a wide evolutionary separation of these groups. 10E. THE EVOLUTION OF THORACIC LEG STRUCTURE IN HEXAPODS The validity of the concept that all arthropodan limbs have been derived from a common basic type of limb, possessing the same number of podomeres, has been questioned and rejected in Part 9, figures 9, 10 and pp on the basis of a comparative study of myriapodan and pterygote classes. While there are sound examples of secondary fusion of leg segments, partial or complete (e.g. coxa with the trochanter of some diplopods and trochanter with the prefemur of some chilopods), and of the converse, a subdivision of leg segments, partial or complete, single or multiple (e.g. the tarsus in certain diplopods, chilopods and pterygotes), it was suggested that such variations have a functional explanation in each case (Parts 6, 8, 9) and that the basic differences in the number of podomeres in the several classes have not arisen by fision or fusion from a common ancestral type. The limb base, the limb tip and the principal knee segment, or one of its joints (providing good flexure at both or only at the distal end), appear to be ' homologous points along the legs of myriapods and hexapods. But the number of joints across the leg, both proximal and distal to the knee, are not similar in each class and the differences are often correlated with known functional needs. Each podomere, therefore, cannot be exactly homologous from class to class, if the subdivisions of a primitive lobopodial leg took place differently in its proximal and distal regions in the various evolving groups of arthropods. No evidence has been found in support of the existence of a hypothetical subcoxa in any apterygote (see above, 0 loc(iii)). This claimed basai segment
; the names of pleurites (embraced by the hypothetical")
167 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 369 of the primitive hexapod leg is accepted by many text-books, e.g. Imms, revised by Richards & Davies (1964), Snodgrass (1952), etc. The name subcoxa has been applied to many apterygote pleurites; to bulges of undifferentiated pleuron (Collembola, Borner, 1921, Denis, 1949); the names of pleurites (embraced by the hypothetical subcoxa) have been applied to parts of the meso- and metathoracic coxae (Collembola, Carpentier, 1949) ; and thickening and folding of part of the proximal rim of the coxa of Thysanura (figure 32D, E), which has no more than mechanical significance, is labelled basicoxite, meaning subcoxa, by Snodgrass (1952, figure 75F) and trochantin by Carpentier (1946). But the subcoxa does not exist in any myriapod or apterygote class (8 7B(iii), 7C(v), loc(iii)). There is more uniformity in the legs of the apterygote hexapods than in the myriapods, apart from their several very different coxa-body junctions. Both coxa and trochanter are mobile and may be regarded as homologous segments. Details of shape and structure of the coxa depend on the various positions of the axis of swing of the coxa on the body, and whether the coxa projects laterally, as in Diplura and Protura, or is pendant, as in Collembola and Thysanura. A rocking movement of the dorsal rim of the coxa takes place only in the Protura and by a different mechanism from that of the Myriapoda (Part 8, figure 3 and p. 258). The coxa executes small (Diplura), considerable (Collembola) and large (Machilidae) adductor-abductor movements in contrast to the myriapod classes and Protura where these movements are almost or entirely absent. The partial and complete coxal subdivisions in Collembola are correlated with functional needs of these animals, in contrast to other classes, and are not primitive. The trochanter segment is very mobile on the coxa in all hexapods, giving good levator movements of the whole telopod. The proximal dicondylic pivot joint is very free, but far less elaborate, than in many myriapods (figures 29, 30, 31, 36). Particular strength and swiftness of movement is provided in Thysanura by the large apodeme projecting into the coxa from the trochanter, bearing depressor muscles, and the non-equatorial position of the pivot articulations (figures 31, 36), such as occur in no other hexapod class. In Collembola and Thysanura, strength, amplitude and probably swiftness of leg movement are promoted by levator muscles which arise, not entirely from the antero-dorsal rim of the trochanter, but from internal tendons anchored on its ventral face, the details differing in the two classes. These features occur in no myriapod or other hexapod class. The distal joint of the trochanter is not very free and proximal trochanter muscles can levate the whole telopod. Unique movements take place at the distal trochanter joint in the Diplura, where some leg rocking provides the same advantage as the coxal rocking of the myriapods. Unique dipluran peculiarities of the femur-tibia joint contribute toward the same movement (5 7A(iii)). The knee flexure is provided by the dicondylic pivot joint at the distal end of the femur in pterygote and apterygote classes. The tarsus is subdivided in respect of the plantigrade stance in the Pterygota. Complete or partial subdivision of the tarsus in certain unguligrade apterygotes, as in some myriapods (Parts 6, p. 503; 8, pp ), serves the needs of fast running and long legs (Thysanura). The leg beyond the trochanter is subdivided into three main segments in
168 370 S. M. MANTON typical members of all hexapod classes, except the Collembola where there are only two. This smaller number is advantageous here in that fewer joints need the internal skeleto-muscular devices which prevent dislocation at a jump by sudden increase in hydrostatic pressure. It is probable that the use of hydrostatic forces by the evolving springing organ was an early characteristic of this class, and that subdivision of the leg, distal to the trochanter, into two segments only is primitive here. It is also probable that other hexapods gained greater flexibility of leg, from a stage with little leg sclerotization, by subdividing the same region into three segments, and that they did not pass through a collembolan-like stage because there was no need for it. The limb tip of hexapods shows two main variants in typical examples of the classes. The plantigrade, multisegmented, tarsus of the Pterygota gives stability over a variety of surfaces, often steeply inclined. Most pterygotes do not execute as long strides as might be expected from the length of their legs, compared with the apterygotes (figure If, g and 8 10B). Their plantigrade stance (in adults) gives secure footfalls to a group in which particular speed is usually obtained by flight. The unguligrade, pointed, tarsus of the apterygote hexapods permits a disposition of footfalls giving greater stride lengths and consequently greater speed of running in exposed situations, but the animals do best on more or less horizontal surfaces. Since the forms of the limb tip, as in the number of ways in which it is possible to use six legs, are limited, the unguligrade condition of the apterygote hexapods may have evolved in parallel, as an alternative to the plantigrade stance of the Pterygota, and in all serves speedy movements. The stability of a multisegmented plantigrade tarsus is also seen in the chilopod Scutigeru (Part 9, figure 10) which can negotiate inclined and vertical surfaces as in the Pterygota. Thus it may be suggested that the leg structure of the hexapods as well as the coxa-body union in the several classes, are so distinct and different one from another, and from the myriapod classes, as to support the view that the legs in each class have evolved independently from a stage when leg structure was starting to differentiate. 10F. THE EVOLUTION OF APTERYGOTE JUMPING Repeatedly jumping high off the ground constitutes an effective escape reaction, making capture difficult by predators. This behaviour is found in Collembola inhabiting the higher levels of soil and litter and in the Thysanura Machilidae. The mechanism of the jump is quite different in the two classes. In both, the structural features involved embrace a large proportion of the whole anatomy and not just that of propulsive organs. By contrast, the specializations of the jumping metathoracic legs of grasshoppers does not render the rest of the body materially different from that found in the less specialized Pterygota. The mechanism of collembolan jumping, described here for the first time (8 9A), shows that the propulsive thrust is largely hydrostatic in origin, aided by the elasticity of cuticular bars. Extrinsic and intrinsic musculature is mainly concerned with folding up the organ after a jump. Intrinsic muscles and the terminal furca gives some control over the jump from uneven surfaces and a minor additional propulsive thrust. The hydrostatic pressure working the jumping-organ is generated by the
169 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 371 trunk musculature, the hamula holding the springing-organ in place until muscular tension and pressure are built up. Rigidity of the body is necessary for the jumping mechanism, and is obtained by cuticular features and by supporting structures. The cuticle everywhere is structurally suited to resist deformation by sudden increases in internal hydrostatic pressure, aided by a number of endoskeletal and muscular devices and by the form of the well-developed superficial pleural musculature. The dorsal and sternal longitudinal systems contain many long sectors traversing several segments of thorax and abdomen, all associated with causing sudden, slight shortening of the trunk, so forcing fluid into the springing-organ. Their action is reinforced by very large deep oblique muscles. A well developed lateral longitudinal system of muscles is present, alone among hexapods, serving the same purpose as the other longitudinals. Specialized sectors of the lateral longitudinals form the main muscles which fold up the springing-organ after use. Deep dorso-ventral muscles form the antagonists to the deep oblique and longitudinal muscles. Thus the trunk musculature is much more specialized than in Diplura, Protura and simple Pterygota, except for the unique retention of the presumably primitive lateral longitudinal muscles (see below). The legs are unique in many features, all of which are correlated with the use of hydrostatic pressure for jumping and the need to prevent leg joints from sliding apart and dislocating at a jump. The coxae have no articulation with the body, a unique condition in a land arthropod as large as Tomocerus. Any of the types of coxa-body articulation of other hexapod classes would be unsuitable here because they are not compatible with an ability to resist the coxae being blown away slightly from the body and dislocated under pressure. Instead the coxa-body union is somewhat like a ball and socket joint, the rim being the flexible membrane surrounding the coxa and the ball being the endoskeletal node situated over the mouth of the coxa. Movements in all direction and twisting on the long axis of the coxa are possible and are caused by very many extrinsic leg muscles. The suspensory mechanism of the leg on the body is unique (figure 30). The usefulness of only two podomeres beyond the trochanter and other details of leg structure have already been noted ( 5 10E). Such an abundance of features associated with jumping can only mean that this habit was one with which the evolutionary differentiation of the class was associated (see further below). The non-jumping Collembola appear to have been derived secondarily from the jumpers in association with living more deeply under cover. Machilid jumping mainly depends upon contrasting features. The evolution of high jumping with the abdomen was probably preceded by the evolution of jumping gaits by the thoracic legs, during which the animal is never quite airborne but supported by only two legs at any one time. Long legs and a good ability to bend the knees, in the sense that knee bending in a vertebrate precedes a good bipedal jump by leg extension, are essential assets of the Machilidae. The very wide adductor-abductor movements of the coxa on the longitudinally elongated pleurite, closely supported by the tergite, are parts of the leg mechanism not found in other classes. Unstable gaits and paired legs moving rapidly in the same phase here can be employed, because the stability of the legs is great enough to take the strain. Leg morphology is suited to these needs (see above).

170 372 S. M. MANTON The abdominal limbs in apterygote hexapods have persisted in recognizable form to a greater extent than in the Pterygota. In all classes these limbs form gonopods posteriorly, but reduced limbs from every or some of the more anterior abdominal segments are present in the apterygote classes and are best developed where the thoracic legs use long strides and unstable gaits. The abdominal limbs appear to act as stabilizers, the styles being dragged over the surface of the ground, muscles holding the style erect (figure 38), see Gzmpodea and Thysanura Pls. 2, figures a, b; 5a, b, c; 6 a-d, where white lines are left by the styles on smoked paper over which the animals had been running. Possibly style dragging may be also sensory. It may be postulated that persistent abdominal limbs in the apterygote hexapods were initially associated with extracting maximum speeds of running out of the thoracic legs, while the Pterygota took an opposite course, one of much greater tarsal stability, useful on non-horizontal surfaces, less speed by thoracic legs, the acquisition of flight and a disappearance of the more anterior abdominal limbs. The evolution of high jumping in the Machilidae presumably followed on a stage when the abdominal limbs were retained for purposes of stability. It would not have been a great step for these limbs then to have been used to push actively against the ground instead of being passively dragged over it. The mobile coxo-sternal plates of the Machilidae could readily have been derived from already small abdominal limbs. Very great specialization of the abdominal musculature, which telescopes the abdominal tergites together dorsally, pressing the posterior free margin of the coxo-sternal plates against the ground, would form the beginning of the present day jumping mechanism. Enlargement of the plates so that they floor two segments, gives more leverage, and the simultaneous use of all plates together gives sufficient thrust to project the animal high into the air. Cuticular features are an important part of the jumping mechanism. The greatly overlapping abdominal tergites, with long paratergal extensions reaching to the ventral level, make the dorsal shortening and ventral expansion of the body a supple movement, yet restricted to the vertical plane. And the thorough boxing-in of the body makes possible the use of hydrostatic as well as muscular forces in pressing the free ends of the coxo-sternal plates against the ground. The cuticular solution of the problems thus differ from those employed by Collembola. The marked size and complexity of the dorsal longitudinal muscles in the abdomen of Petrobius, together with the presence of multiple sectors of the deep oblique muscles, and their spread to several tergites dorsally from the one point of origin (figures 38, 39), all serve the sudden dorsal pulling together of the abdominal tergites. These functions are in keeping with those of the same categories of muscles in other hexapods and in myriapods. The deep dorso-ventrals act as antagonists. The Collembola meet the needs of body shortening by simple enlargement of the deep dorso-ventral and deep oblique muscles instead of the machilid multiplication of sectors in each category. Clearly the two musculatures have evolved independently. Special machilid features are the simple segmental series of ventral or sternal longitudinal muscles, used for the recovery movement of straightening the abdomen after a jump. The contrasting elaboration of this category of muscles in Collembola is associated with their use, not as antagonists of the dorsal longitudinals and the

171 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 373 deep muscles, but as contributors to the general body shortening, produced by the dorsal and lateral longitudinals and the deep muscles. There are other peculiarities of machilid musculature whose functions are not fully understood. Striking similarities in some details are shared with the several malacostracan orders which jump backwards through the water by sudden flexure of the abdomen. In these Malacostraca and in Petrobius the length of the dorsal muscles is increased by their being twisted round one another, and in both the deep oblique musculature is looped. In Petrobius two of these muscles (25B, 25A) pass from the ventro-lateral tendons up to the dorsal region where they loop round two other sectors (24, 26) and then descend to the tergite behind and to a following segmental tendon respectively. Roughly similar, but more complex, looping is found in crustacean abdomens in association with jumping through the water. It is suggested here that displacements of the exoskeleton may be increased or facilitated by the presence of rope-like and looped muscles. Whether elastic forces are particularly involved is not known. In these Malacostraca and in Machilidae there is no mechanism for holding the springing organs in place while muscular tension builds up, as there is in Collembola, by the hamula, and jumping dipterous larvae, by the jaws. Clearly the field is wide open for physiologists to study the problems involved ( 5 9D). Thus the jumping mechanisms of the Collembola and Thysanura show two entirely different modes of meeting a common need, that of gaining protection when venturing into exposed situations. Flight is an impossible evolutionary line for all the apterygote classes because their coxa-body junction and pleural arrangements are unsuitable. Instead, these diverse jumping mechanisms have been evolved. In the Collembola the essay must have begun along with the differentiation of the class, since its perfection involves almost the whole anatomy. In the Thysanura the morphological features associated with jumping gaits are limited to the thorax and those associated with high jumping mainly to the abdomen. 10G. THE EVOLUTION OF HEXAPOD CLASSES A comparative and functional survey of the modes of action of the jaws or mandibles in the major groups of arthropods (Manton, 1964) has shown how decisive are the similarities and differences between the several groups. The Onychophora, Myriapoda and Hexapoda have in common a mandible which is a whole limb, biting with the tip, in contrast to the Crustacea, Limulus and arachnids where biting or grinding is done by a gnathobase. It was concluded that the evolution of the Onychophora-Myriapoda-Hexapoda did not proceed from any of the known basically aquatic groups, but from an independent, presumable aquatic, stock developing its own solution of the need for a mandible. The further differentiation of these three terrestrial groups must then have proceeded in parallel. The Hexapoda have in common a head capsule with an unjointed mandible, primitively moving by a rolling action comparable to the promotor-remotor movement of an ambulatory limb. This movement provides grinding by molar surfaces. Biting in the transverse plane is a secondary derivative from the primitive rolling mandible, whose movement is seen very simply in Petrobius

172 3 74 S. M. MANTON today. But the Machilidae have improved on the usefulness of a purely grinding mandible by the formation of distal scraping tips, far removed from the molar processes, and a hydraulic mechanism for raising the particles (unicellular algae) to the grinding molar surfaces. Hexapods also have in common the same number of post-oral feeding limbs, including a labium. The Hexapoda possess anterior and posterior tentorial apodemes in the head. The anterior tentorial apodemes are fairly rigid, and in hexapods employing the primitive rolling mandibular movements, carry the principal extrinsic mandibular muscles (e.g. Petrobius, Manton, 1964, Fig. 21). The evolution of these apodemes is presumably associated with the hexapod type of mandible. The transition towards more specialized mandibular movements in hexapods brings a shift of the principal mandibular extrinsic muscles to other sites (e.g. locust, loc.cit., Fig. 29) or the mandibular mechanism may be such that these apodemes are not required (e.g. Cumpodeu where they are absent, 1oc.cit.). The posterior tentorial apodemes are associated basically with the provision of suitable sites for the insertion of extrinsic labial muscles in hexapods (e.g. Petrobius, Ioc.cit., Fig. 24J), a function retained by these apodemes, amid many diverse specializations, in Ctenolepisma, locust, Tomocerus, Campodea, etc. (as shown by Figs. 27, 29, 35, 47 1oc.cit.). The extrinsic muscles from maxilla 1 insert on either or both of the two pairs of apodemes as is spatially convenient. The anterior and posterior tentorial apodemes show stages of fusion along with the development of secondary biting in the transverse plane in the Lepismatidae, and the process of fusion is complete in the Pterygota, correlated with stronger transverse biting and a need for a strongly supported head capsule carrying the mandibular adductor muscles (Fig. 29, 1oc.cit.). The pterygote tentorium, formed in this manner, supports also the strongly musculated labium with a large area of free anterior face; this face is small on the symphylan maxilla 2 and diplopod gnathochilarium. The two pairs of tentorial apodemes in the entognathous apterygotes remain separate because of the development of protrusible mandibles. Thus a fairly rigid anterior tentorial apodeme is associated with the basic type of hexapod mandible and the posterior tentorial apodeme is functionally associated with the hexapod labium; all these structures must have characterized the ancestors of the hexapod classes. The Myriapoda contrast in a basic possession of a jointed mandible with primitive biting in the transverse plane. Mandibular adduction is caused by mandibular muscles, but abduction is mainly effected by a push delivered on each mandible by a mobile, swinging anterior tentorial apodeme. No posterior tentorial apodemes are present. Maxilla 2 of the Symphyla and the gnathochilarium of the Diplopoda have some functional resemblances with the hexapod labium. Maxilla 2 muscles in Scutigerellu insert on the exceptionally long and posteriorly-elongated anterior tentorial apodeme (Fig. 58b, 1oc.cit.). But only late during development is the symphylan maxilla 2 segment added to the head from the trunk (Tiegs, 1940). Neither the posterior tentorial apodeme nor an associated typical labium is present in any myriapod. The Onychophora possess an unjointed very short mandible, moving just as do the walking legs, cutting food with an antero-posterior slicing by jaw blades which are comparable with enlarged paired claws of the walking legs. No head apodemes or transverse tendons are present, they would be incompatible with the habit of basic importance, that of extreme ability to alter the body shape

173 EVOLUTION OF HEXAFQD LOCOMOTORY MECHANISMS 375 and squeeze through crevices without pushing. Each jaw has a longitudinally situated apodeme. An ability to form well sclerotized structures in the head is thus in evidence in the jaw blades and jaw apodemes. The Hexapoda, Myriapoda and Onychophora are sharply distinct from each other in their jaw anatomy, jaw mechanisms and head endoskeleton, although sharing a type of jaw or mandible which contrasts with those of all other arthropods (Manton, 1964). Hexapod unity, shown by their mandibles and head endoskeleton, is also seen in the embryonic configuration of the head which is identical in all hexapod classes, in contrast to onychophoran and myriapodan heads (Anderson, in press). But the unity of the Hexapoda indicated by head structure, although real, must be a distant one because each of the five component classes shown such contrasting features: the mutually exclusive modes of union of the coxa with the body and the axis of coxal movements; the external and internal thoracic structure, including the pleural armature, trunk endoskeleton and muscles; the differences in the thoracic limbs and in the gaits used; and all these features meet the variety of needs. These systems are so different one from another (5 10A-F) as to indicate that nothing less than class status is suitable for the five groups of hexapods. Musculature is plastic to a considerable degree, one or both ends of a muscle can shift according to needs and even move out of its initial segment (see examples in Part 7, 5 19, Part 8, 5 8v and figure 34). But leg and tentorial mechanisms dependent upon cuticular skeleton show no signs of an ability to change towards one or another existing type, and it is difficult to see that any one of the myriapod or hexapod coxa-body junctions and the movements they support, could have entirely changed their characteristics while preserving functional continuity. The five hexapod classes thus appear to result from independent and parallel modes of evolution. The Collembola have been shown above to be very far removed from a primitive hexapod state. No other class exhibits such extreme morphological modifications of the whole body associated with hydrostatic jumping. The ancestors of Collembola have probably been distinct from those of other hexapods since a stage with a soft-bodied trunk in which hydrostatic forces played a great part, as in Onychophora today (Manton, 1958c, 1967). A differentiation of anterior limbs for walking and posterior limbs for jumping would be an easy step for a primitive lobopodial animal, or from one in which cuticular stiffening was starting. Onychophora often use paired legs in similar phase and progressing at their fastest have few legs in contact with the ground (Part 1, P1. 15, figure 12). A subterminal pair of legs on a soft bodied collembolan ancestor might easily have pushed in unison and initiated a jumping movement. No class, other than Collembola, has bound itself to hydrostatic trunk rigidity, the collembolan type of jumping and associated morphological changes, including a retention of the lateral longitudinal muscular system. And the retention of these features and the high specialization of others does not lead towards any other class of hexapod. The Diplura contrast with the Collembola, both functionally and anatomically, in all the systems reviewed above in The distinction between the dipluran coxa-body junction and the support thereof and the corresponding features of Collembola and other hexapods, suggests that only parallel evolution from ancestors with a soft body could account for these features. The Diplura show some superficial thoracic similarities with
174 376 S. M. MANTON myriapods and Protura, but the details contrast, and limbbases and limb-rocking mechanisms, must be the result of independent evolution. The Thysanura comprise two groups built upon a remarkably common plan, showing similar, unique features of thoracic leg construction and action, similar types of coxa-body union, tendinous and apodemal endoskeleton, abdominal limbs etc. But each family is highly adapted for certain performances. The Machilidae preserve a primitive rolling mandible, but the head is specialized for sucking up algae (see above), and the thorax and abdomen are highly specialized for the performance of jumping gaits and abdominal jumping escape reactions. The Lepismatidae have advanced towards transverse biting, with associated changes of head structure (Manton, 1964), they have their own highly specialized use of the legs and jumping gaits, which probably depended upon an initial condition resembling the essentials of machilid legs. The two families are divergent. But because the Lepismatidae have progressed in the direction of strong, hinged, transversely biting mandibles with a wider gape, although much less far than have the Pterygota, the Lepismatidae should not be regarded as showing greater affinity with the Pterygota than do the Machilidae. The lepismatid similarities with the pterygotes are convergent and do not indicate affinity. Neither do the Machilidae show a number of characters in common with the Symphyla (Heeg, 1967). The Machilidae in fact show such fundamental divergence from the Symphyla as to preclude relationship, as shown by their entirely different leg and jaw mechanisms which could not have evolved the one from the other (see Part 9). The Thysanura in no way occupy an intermediate position between Pterygota and Symphyla and exhibit fundamental differences from all myriapod classes in their head structure (Manton, 1964), limbs, pleuron, coxa-body junction, etc., as shown here. Thysanura possess a coxa-body junction with a pleurite, as in Pterygota, but the pleurite is mobile in Thysanura and fixed in Pterygora and could not function otherwise in either class. The pleurites of Lepisma do not show a primitive or basic condition which could have led to pleural morphology of other classes (see above). Rigidity of the principal pleurite in the Pterygota was a prerequisite for the evolution of flight. The Pterygota have shown no tendency towards the machilid type of jumping gaits or high jumping with its extreme abdominal specializations, nor have the Pterygota adopted the remarkable leg movement of the Lepismatidae. In both Machilidae and Lepismatidae much associated morphology facilitates the proficiencies. The Thysanura and Pterygota must have been divergent from a very early stage. The Protura, under 2 mm in length, lacking antennae and possessing entognathy in the head, with a quadrupedal mode of progression and skeleto-muscular features conferring trunk flexibility, do not suggest close affinity with any other class. The presence of a well-sclerotized sternite as well as tergite, a wide pleuron with many pleurites, a coxa-sternite articulation, a leg-rocking mechanism and trunk musculature conferring mobility rather than rigidity are parallel evolutions in Myriapoda, Diplura and Protura because the details are so different. The proturan coxa articulating with both a pleurite and a sternite is unique among hexapods, as is the associated coxa-pleural rocking mechanism.
175 EVOLUTION OF HEXAPOD MCOMOTORY MECHANISMS 377 Tuxen (1970) reiterates his belief in the unity of the Entognatha linking together the Collembola, Diplura and Protura. Yet he does not produce any evidence to contradict the mass of reasoned morphological and functional detail concerning this matter put forward by Manton (1964). The essential asset conferred by entognathy is a protrusible mandible, one which is very freely associated with the head in contrast to a strongly-hinged mandible used for transverse biting. A mass of morphological features enable mandibles to be protrusible, including muscles causing this action. A boxing-in of a weakly attached mandible is also essential, and is done in different manners by the various examples. Entognathy and mandibular protrusion is a widespread phenomenon, occurring in Pauropoda, Chilopoda, siphonophoran Diplopoda, Onychophora, many Crustacea etc. The mechanisms of mandibular protrusion differ in the non-hexapod examples, and they differ in the entognathous hexapods. The differences between collembolan and dipluran entognathy have been listed in detail by Manton (1964, pp. 65, 70, etc.), where it was concluded that they were so great as to indicate parallel evolution of entognathy in the two classes. In view of the additional data concerning the trunk recorded above, any supposed community of origin of Collembola and Diplura is quite untenable. The former possess an anatomy geared entirely towards hydrostatic jumping and trunk rigidity, while the Diplura show quite other trunk features and accomplishments, conferring flexibility and not rigidity, and show entirely different types of endoskeleton, leg support and leg movements. Even the entognathy of the Collembola is associated with remarkable sheets of connective tissue and tough membranes which box-in the salivary glands and other structures in the head and presumably protect them from sudden pressure changes (Manton, 1964, Figs. 32, 3 3, 40P, posterior endoskeletal plate and unlabelled dorso-ventral membrane just internal to the salivary ducts on figure 40P). The significance of these structures was not appreciated at the time when they were figured, owing to the jumping mechanism and use of hydrostatic forces being then unknown. No such pressure shields exist in the dipluran head. Although the proturan head anatomy has not been worked out in a comparable comparative and functional manner as for Collembola and Diplura, the overall peculiarities of the class, noted above, do not suggest any affinity with the Collembola or with the Diplura. If the Entognatha are dismissed as a convergent assemblage in which protrusible mandibles have been evolved, along with many contrasting features, what of the Ectognatha comprising the Monura, Thysanura and Pterygota? The present work and that on the heads of arthropods (Manton, 1964) suggests that there can be no common ancestry which particularly links the Thysanura and Pterygota, because the five hexapod groups possess mutally exclusive leg mechanisms, although sharing the same basic type of jaw mechanism which contrasts with all myriapods. The Thysanura and Pterygota are essentially divergent classes. If the taxon Entognatha be dissolved as invalid, as suggested (Manton, 1964) and reinforced by further evidence here, there is little purpose in retaining the invalid term Ectognatha. Thus hexapod evolution may be envisaged as stemming from one of Sedgwick s (1909) hypothetical and varied primitive land arthropods, which we
176 378 S. M. MANTON can now define more exactly. Such arthropods at their earliest appearance on land probably possessed many lobopodial limbs, little sclerotization and a habit of dealing with their food by the tips of some anterior limbs. Jaws or mandibles could have been evolved from such limbs, and on the second body-segment in the archi-onychophora but on the fourth segment in others giving extant descendants. The exact segment on which special organs are situated is often freely alterable; the clasping organs in Branchiopoda arise on a variety of anterior segments in different species, and the powerful feeding clasper in some harpacticids (e.g. Amphiascus), which is first situated on the antenna1 segment and later on the maxillipedal segment in the life history of the same animal. The habit of predominant importance to the Onychophora, that of extreme distortability of body, must have been established early and led to the evolution of the onychophoran head and trunk characteristics (Manton, 1958c, 1967). In other early land arthropods two types of encapsulated heads arose, the archi-myriapods with jointed transversely biting mandibles and one pair of tentorial apodemes and the archi-hexapods with rolling, grinding mandibles, and two pairs of tentorial apodemes and a labium. The body in both was probably little sclerotized and the limbs many and lobopodial. Since functional continuity must have been maintained through all evolutionary stages, and since the leg mechanisms of the hexapod classes are mutually exclusive, these classes cannot have given rise one to another and must have evolved in parallel from ancestors with lobopodial limbs and little trunk sclerotization. Incipient thoracic limbs, differentiated from abdominal ones, could increase in size only when trunk and limb sclerotization developed. The differing coxa-body unions and leg mechanisms of the hexapod classes must have evolved in parallel and early differentiations, both of habits and of uses of legs, must have been important. The ancestors of the hexapod classes thus could not have been hexapodous, but multilegged. Phrases such as the ancestral hexapod (Heeg, 1967) become meaningless because there never could have been such an animal. As sclerotization and the differing leg mechanisms became established, so the three thoracic segments presumably became differentiated from the rest. It must be concluded that the hexapodous state and the formation of a thorax are a parallel evolution in the five hexapod classes. Since a hexapodous use of legs is found in Galeodes at all times, in spiders sometimes, in some prawns, some crabs etc. there is nothing improbable in the parallel evolution of a hexapodous thorax in the five pterygote and apterygote classes. The many theories of origin of the Pterygota, ranging in postulated descent from Crustacea to Myriapoda, Symphyla and Apterygota, can now be set aside, because none of them provide functional continuity in the evolution of essential morphology. Brauer (1869) claims descent of the Pterygota from Onychophora, via Chilopoda and Campodeidae; Hansen (1930) from Syncarida; Crampton (1928) from Isopoda; Verluys and Detmoll (1921) from Onychophora via the Myriapoda; Handlirsch ( 1908) from Trilobita; Tillyard (1930) joint ancestry of Pterygota and Myriapoda from an unknown stem, placing Collembola at the base of the insectan series; Calman (1936) from ancestors much as in existing Thysanura, the Symphyla giving hints as to their immediate predecessors; Imms (1937) supports a common origin of Symphyla
177 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS and Thysanura, claiming in his textbook (1964) that the most primitive insects are to be found in the Thysanura and Diplura ; while Hughes (1965) categorically states that insects evolved from a myriapodan stock. All these views fail because, on proper examination, a functionally impossible evolutionary series of animals is postulated by each. The comparative study of functional morphology in arthropods, recorded in Parts 1-10 has been rich in the information it has yielded on the past history of these animals. This study, with that on the head, embraces the whole anatomy, excluding digestive, reproductive and nervous systems and, correlated with knowledge of locomotory and other habits, has given the conclusions set out in figure 40, which differ so greatly from those of all other schemes, based upon inadequate and non-functional information. Tiegs ( 1947) put forward the evidence of relationship between the Onychophora, Myriapoda and Hexapoda, a conclusion amply substantiated by the present work and by the embryological studies of Anderson (in press). This implies the independent acquisition of jointed legs and trunk sclerotization by these land animals and by the aquatic Crustacea, Chelicerata, Trilobita etc., bringing with it the inescapable conclusion that the Arthropoda are polyphyletic (see further in Manton, 1964). But so far we have little hint as to what the soft-bodied ancestors of the sclerotized aquatic arthropods were like. Terrestrial Crustacea and Arachnida must have possessed sclerites before they reached the land, hut the aquatic ancestors of the Onychophora-Myriapoda-Hexapoda assemblage probably left the sea with soft bodies and lobopodial limbs. The relatedness of the Onychophora-Myriapoda-Hexapoda assemblage of animals is now beyond question. This cumbersome form of reference has been in use for some years. It is here proposed that these three groups be termed the Uniramia, arthropods with primitively uniramous limbs, in contrast to the basically biramous-limbed Crustacea, Trilobita and Chelicerata, in which the uniramous condition, when it occurs, is secondary. The erection of the taxon Uniramia does not imply that all biramous limbs are basically similar-indeed they are not. Secondly the Uniramia have in common a jaw or mandible which is a whole limb, biting with the tip, mlike all other Arthropoda. The Crustacea contrast with their mandibular gnathobase, the primitive movement of which is a roll, giving grinding, unlike the Chelicerata where the primitive movement is chewing (biting) in the transverse plane. Many advanced Crustacea bite in the transverse plane, but this is a secondary achievement derived from the more primitive rolling mandible (Manton, 1964). Thirdly the Uniramia have in common a long mid-gut, food being retained in particulate form in the lumen of the tube. In the Crustacea the midgut is frequently very short, and in both Crustacea and Chelicerata food particles pass from the gut to diverticula in which digestion and sometimes absorption take place. Fourthly, the Uniramia have in common embryological characteristics which are not present in Crustacea and Chelicerata (Anderson, 1969, in press). It seems fitting to give the status of phylum to the Uniramia now that the Arthropoda can no longer be regarded as a monophyletic group. The Uniramia appear to have developed their cuticular armour of sclerites independently of the other Arthropoda and to have done this much more recently. How many arthropodan phyla there may be in all is uncertain, but among extant 379
178 TIME soft, multilegged body. lobopodial limbs, manipulating food (incipient biting) by the tips of future jaws, contrasting with Chelicerata and Crustacea, each of which possess a different type of sclerotized gnathobasic jaw and different type of biramous limb > n No head capsule. sclerotized jaws and jaw apodeme present, multilegged soft trunk, TIME > > ONYCHOPHORA MYRIAPODA DIPLOPODA -- 3 > PAUROPODA CHILOPODA ) SYMPHYLA 3 % \ soft, multilegged body, HEXAPODA.- Sclerotized trunk. Sclerotized head capsule, unjointed. rolling mandibles, 2 pairs tentorial apodemes. THYSANURA 3 and thorax in each class. labium, multilegged trunk. lobopodial limbs PTERYGOTA > Figure 40. Diagrammatic representation of the conclusions reached concerning the evolution of the Uniramia (comprising the Onychophora! Myriapoda, Hexapoda, with primitively uniramous limbs) from soft bodied, multilegged ancestors which were distinct from the ancestors of the Crustacea and Chellcerata, tlme progressing from left to right. These conclusions are based upon the evidence of functional morphology presented here and upon that concerning the jaws or mandibles (Manton, 1964). Speculative dichotomies have been omitted. As yet there is no evidence which determines with certainty whether the Dignatha (Pauropoda, Diplopoda) forms a grade or a taxon, but it is probably a taxon (see text). The Trignatha (the hexapod classes and the Symphyla) is certainly a grade and not a taxon. For further description see text. W m 0 f

179 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANlSMS 381 arthropods the Crustacea and Chelicerata contrast in status with the Uniramia. Within the Uniramia it is here proposed that the three groups: Onychophora, Myriapoda and Hexapoda be each accorded the rank of subphylum. The Arthropoda must now be regarded as a grade of advancement, similar to those of mammals and reptiles, which has been reached independently several times by metamerically segmented animals such as Trilobita, Chelicerata, Crustacea and Uniramia. Doubt has been cast on the use of the terms Monognatha, Dignatha and Trignatha as taxonomic categories, it being more probable that they represent grades of organization within the Uniramia (Manton, 1964). Some suggestions, emerging from figure 40 and from the various functional analyses of morphological systems, can now be made concerning these grades and with particular reference to (i) a supposed evolutionary dichotomy ending in the Pauropoda and Diplopoda (the Dignatha in the older taxonomic sense) and (ii) the evolution of the Trignatha. There seem to be no valid reasons for supposing that a dignathan state was preceded phylogenetically by a monognathan one or that hexapod trignathy was superimposed upon an ancestral dignathan head. These three types of head represent different ways of meeting a common need and trignathy has been evolved several times. All Myriapoda possess a functional and highly integrated unit comprising: a particular type of mandible, a swinging anterior tentorial apodeme and a maxilla, all with appropriate musculature, connections and movements. It is possible to regard all the myriapod classes as basically dignathan, with varied posterior additions to the head in some classes, in which the additions lack both structural and functional uniformity. The Pauropoda show no additions to the basic dignathan head. The Diplopoda possess a post-gnathochilarial segment without limbs which, embryologically, contributes both to the posterior part of the head and to the collum segment (Dohle, 1964). In the Chilopoda a maxilla 2 segment, with limbs, forms part of the head behind the basic dignathan unit, the limbs, as mouth parts, are little modified. The Symphyla also possess a maxilla 2 segment, which starts its embryonic development as part of the trunk and later in ontogeny becomes united with the head. But this is no typical labial segment, such as occurs in the trignathan Hexapoda: there are no posterior tentorial apodemes and resemblances to a labium are partial and convergent (Manton, 1964). The trignathy of Chilopoda and Symphyla is superimposed upon basic myriapodan dignathy. The diplopod condition has not advanced so far as in the Symphyla, in that only part of the post-gnathochilarial segment has been incorporated into the head. But the Chilopoda have progressed beyond a trignathan state, in that the poison claw segment functionally forms part of the head. Thus it appears to be more suitable to regard the whole of the Myriapoda as basically dignathan, with various additional head features of a diverse nature occurring in the several classes, than to consider the Pauropoda and Diplopoda as two divergent branches of a single dignathan stem. The trignathan heads of the hexapod classes contrast with those of the Myriapoda in that hexapod trignathy is based upon a quite different functional unit comprising: a particular type of mandible requiring fixed apodemal support, maxilla 1, labium and two pairs of fixed tentorial apodemes. This more elaborate unit cannot have arisen by fundamental alterations of the basic
180 382 S. M. MANTON dignathan unit and addition of a maxilla 2 segment with posterior tentorial apodemes. The well integrated hexapod trignathy has been very flexible in its evolutionary adaptations, but cannot be regarded as having evolved from a dignathan state as has the trignathy of Chilopoda and Symphyla. Hexapod heads have probably been trignathan as far back as their earliest differentiation. Thus the basic functional dignathan unit of all Myriapoda and the trignathan unit of Hexapoda are entirely distinct, the one could not have arisen from the other but both could be descended from early arthropods showing different patterns of cephalization. Similarly a monognathan jaw unit, such as seen in the Onychophora, cannot have been a precursor of the dignathan or trignathan units because the differences between them are too great. ACKNOWLEDGEMENTS I wish to thank the many persons who have kindly supplied me with living animals, in particular: Dr H. Goto for cultures of Collembola, Mr and Mrs Greenslade for japygids from the Solomon Islands, Mr J. S. CoIman for Petrobius from Port Erin and the Staff of the Marine Biological Association at Plymouth. Dr T. Clay and other members of the British Museum, Entomology Department have kindly identified specimens for me. Dr J. P. Harding has rendered his usual assistance with the photography and Mr J. G. Blower and Professor D. T. Anderson have kindly read the manuscript with care and made many valuable suggestions. I also thank the British Museum (Natural History) for hospitality during the execution of this work and for access to preserved material. I gratefully acknowledge the financial assistance given to me by the Science Research Council. REFERENCES ANDERSON, D. T., The comparative early embryology of the Oligochaeta, Hirundinea and Onychophora. Proc. Linn. SOC. N.S. W.. 91: 10-43, 68 text-figs. ANDERSON, D. T., On the embryology of the cirrepede crustaceans Tetraclita rosea (Krauss), Tetmclita purpurascens (Wood), Chthalmus anrennarus Darwin and Chamaesipho columnar (Spengler), and some considerations of arthropod phylogeny. Phil. Trans. R. SOC. (B), 256: , 30 text-figs. ANDERSON, D. T., In press. Embryology and phylogeny of Annelids and Arthropods. Pergamon Press BARLET, J., Remarques sur la musculature thoracique des Machilides (Insectes Thysanoures). Annls Soc. scient. Brux., 60: 77-84, 2 text-figs. BARLET, J., La question des pieces pleurales du thorax des Machilides (Thysanoures). Bull. Annls SOC. r. ent. Belg., 86: , 3 text-figs. BARLET, J., Morphologie du thorax de Lepisma saccharina L. (AptCrygote Thysanoure). Bull. Annls SOC. r. ent. Belg., 87: , 1 text-fig. BARLET, J., Morphologie du thorax de Lepisma sacchnrina L. (Aptkrygote Thysanoure) I1 Musculature(1re partie). Bull. Annls Soc. r. ent. Belg., 89: , 2 text-figs. BARLET, J., Morphologie du thorax de Lepisma saccharina L. (AptCrygote Thysanoure) 111 Musculature (2me partie). Bull. Annls SOC. r. enr. Belg., 90: , 2 text-figs. BARLET, J., Squelette et musculature thoraciques de Lepismachilis Y-signata Kratochvil (Thysanoures). Bull. Annls SOC. r. ent. Belg., I03: , 3 text-figs. BARLET, J. & CARPENTIER, F., Le thorax des Japygides. Bull. Annls SOC. r. enr. Belg., 98: , 2 text-figs. BERLESE, A., Monografia dei Myrientomata. Redia VI (1909); 1-182, 13 text-figs., 17 pls. BORNER, C., Die Gliederung der Laufbeine der Atelocerata Heymons, Sber. Ces. Naturforch, 1902: , 2 PlS. BORNER, C., Die Gliedmassen der Arthropoden. In Lang s Handbuch der Morphologie der wirbellosen Tiers. Jena: , 57 text-figs. BRAUER, F., Betrachtungen ii.d. Verwandlung der Inseckten imm Sinne der Descendenz-Theorie. Verh bot. Ges. Wien, XIX: , 1 pl.
. Bull. Annls Sor. r. ent. Eelg., 82: 165-181, 6 text-figs. CARPENTIER, F., 1949.")
181 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 383 CALMAN, W. T., The origin of insects. Proc. Linn. SOC. Lond.. ( ): , 5 text-figs. CARPENTIER, F Sur la valeur morphologique des pleurites du thorax des Machilides (Thysanoures). Bull. Annls Sor. r. ent. Eelg., 82: , 6 text-figs. CARPENTIER, F., A propos des endosternites du thorax des Collemboles (Apterygotes). Bull. Annls. SOC. r. ent. Belg., 85: 41-52, 5 text-figs. CARPENTIER, F., Pleurites thoraciques de Lepisme et pleurites de Blatte. Bull. Annls. SOC. r. ent. Belg., 91: , 1 text-fig. CARPENTIER, F. & BARLET, J., Les Sclkrites pleuraux du thorax de Campodea. Bull. Inst. r. Sci. nat. Belg., 27: 1-7. CARPENTIER, F. & BARLET, J., The first leg segments in the Crustacea Malacostraca and the Insects. Smithson. misc. Collns, 137: CRAMPTON, G. C., The evolution of insects, chilopods, diplopods, Crustacea, and other arthropods indicated by a study of the head capsule. Can. Ent.. 60: , pls DANIEL, R. J., The abdominal muscular systems of Praunus flexuosus (Miiller). Rep. Lancs. Sea-Fish. Labs, 1927: 5-41, 1 pl. DANIEL, R. J., The abdominal muscular systems of Meganyctiphanes norvegica (M. Sars). Rep. Lancs. Sea-Fish. Labs, 1928: , 1 pl. DANIEL, R. J., Comparative study of the abdominal musculature in Malacostraca. Part 1. The main ventral muscles of the typical abdominal segments. Rep. Lancs. Sea-Fish. Labs, 1930: 57-71, 2 pls. DANIEL, R. J., The abdominal muscular systems of Homarus vulgaris (L.) and Palinurns vulgaris (Latr.). Rep. Lancs. Sea-Fish. Labs. 1930: 3-48, 2 pls. DANIEL, R. J., The abdominal muscular systems of Paranaspides lacustris (Smith). Rep. Lams. Sea-Fish. Labs, 1931: 26-44, 1 pl. DANIEL, R. J., Comparative study of the abdominal musculature in Malacostraca. Part 11. The superficial and main ventral muscles, and their continuation into the thorax. Rep. Lancs. Sea-Fish Labs, 1931: , 3 PIS. DANIELS, R. J., Comparative study of the abdominal muscular systems of Lophogaster typicus, M. Sars, and Gnathophausia zoea, Suhrn, and their relationships with the musculatures of other Malacostraca. Rep. Lancs. Sea-Fish. Labs, 1932: , 2 PIS. DENIS, R., SousClasse des Apterygotes. In Grass6 (Ed.), Trail; de Zoologie, 9: , 114 text-figs. DOHLE, W., Die Embryonalentwicklung von Glomeris marginata (Villiers) irn Vergleich zur Entwicklung anderer Diplopoden. Zool. Jb. Anat. Ont., 81: DRIFT, J. VAN DER, Analysis of the animal community in a beech forest floor. Tijdschr. Ent., 94: 1-168, 16 text-figs., 1 pl., 3 tables. EVANS, M. E. G., The muscular and reproductive systems of Atomariu Ruficornis (Marsham) (Coleoptera, Cryptophagidae). Trans. R. SOC. Edinb., 64: , 64 text-figs. FERRIS, G. F., The myth of the thoracic sternites in insects. Microentomology, 5: GOUIN, F. T., Le thorax imaginal des insectes B la lumiere des travaux rkents. Annls. bid, 35: , 11 text-figs., 4 tables. GRAY, J., 1968, Animal locomotion. i-xi, London: Weidenfeld & Nicolson. HANDLIRSCH, A,, Die fossilen lnsecten und die Phylogenie der rezenten Formen. Handb. fur Palaeont, und Zool., 8-9 Engelmann: , 3 pls. HANDSCHIN, E., Die Okologie der Collembolenfauna westfalischer Hochmoore. 2. wiss. lnst. Eiol., 22: , 3 tables. HANSEN, H. J., Studies on Arthropoda. IIl. On the comparative morphology of the appendages in the Arthropoda. E, Crustacea (supplement), Insects, Myriapoda, and Arachnida: 1-376, 16 PIS. Copenhagen. HEEG, J., Studies on Thysanura, 1, The water economy of Machiloides delanyi Wygodzinsky and Ctenolepisma longicaudata Escherich. Zoologica Africana 3(1) 21-41, 8 tables, 7 text-figs. HOLST, E. VON, Uber die Ordnung and Umordnung der Beinbewegungen bei Hundertfiissern (Chilopoden). Pfugers. Arch. ges. Physiol.. 234: , 7 text-figs. HOWELL, A. B., Speed in animals. their specialization for running and leaping, i-xii, 1-270, 55 text-figs. University of Chicago Press. HUGHES, G. M., The coordination of insect movements. 1, The walking movements of insects. J. exp. Biol.. 29: , 8 text-figs., 1 pl. HUGHES, G. M., Locomotion: terrestrial. The physiology of insects, 2: , 16 text-figs. New York: Academic Press. IMMS, A. D., The ancestry of insects. Nature, Lond., 139: 399. IMMS, A. D., On the antenna1 musculature in insects and other arthropods. Q. Jl microsc. Sci. Lond., 81: , 25 text-figs. IMMS, A. D., A general textbook of entomology, (revised by 0. W. Richards & R. G. Davies); i-x, 1-886, 609 text-figs. JEANNEL, R., Classification et Phylogenie. Les Insectes Fossiles. Evolution et Geonomie. In P. Grass.6, (Ed.), Trait6 de Zoologie, 9: 1-110, 109 text-figs.
182 3 84 S. M. MANTON KUHNELT, W., Soil biology, 1-397, 77 text-figs., 4 pls. London: LUBBOCK, J., Monograph of the Collembola and Thysanura. Ray Soc. Monogr., 1871: i-x, 1-276, 78 pls. McCONNELL, E. & RICHARDS, A. GLENN, How fast can a cockroach run? Bull. Brooklyn ent. SOC., 50: 36-43, 3 text-figs. MAKI, T., Studies on the thoracic musculature of insects. Mem. Fac. Sci. Taihoku imp. Univ., 24: 1-343, 17 PIS. MANTON, S. M., On the emhyology of the mysid crustacean, Hemimysis lamornae. Phil. Trans. R. SOC. pi, 216: , 4 pls., 32 text-figs. MANTON, S. M., On the embryology of the crustacean Nebalia bipes. Phil. Trans. R. SOC. (B). 223; , 6 pls., 17 text-figs. MANTON, S. M., The evolution of arthropodan locomotory mechanisms. Part 1. The locomotion of Peripatus. J. Linn. SOC. (Zool.), 41: , 9 text-figs., 4 PIS. MANTON, S. M., 1952a. Part 2. Geneneral introduction to the locomotory mechanisms of the Arthropoda. J. Linn. SOC. (Zool.). 42: , 5 text-figs. MANTON, S. M., 1952b. Part 3. The locomotion of the Chilopoda and Pauropoda. J. Linn. SOC. (Zool.), 42: , 12 text-figs., 6 pls. MANTON, S. M., Part4. The structure, habits and evolution of the Diplopoda. J. Linn. SOC. (Zool.), 42: , 8 text-figs., 4 pls. MANTON, S. M., Part 5. The structure, habits and evolution of the Pselaphognatha (Diplopoda). J. Linn. SOC. (Zool.), 43: , 8 text-figs., 1 pl. MANTON, S. M. 1958a. Hydrostatic pressure and leg extension in arrhropods with special reference to arachnids. Ann. Mag. nut. Hist. (Ser. 131, 1: , 5 text-figs., 1 pl. MANTON, S. M., 1958b. The evolution of arthropodan locomotory mechanisms. Part 6. Habits and evolution of the Lysiopetaloidea (Diplopoda), some principles of the leg design in Diplopoda and Chilopoda, and limb structure in Diplopoda. J. Linn. Soc. (Zool.), 43; , 21 text-figs., 1 PI. MANTON, S. M., 1958~. Habits of life and evolution of body design in Arthropoda. J. Linn. SOC. (Zool.), 44: 58-72, 1 text-fig., 2 pls. MANTON, S. M., The evolution of arthropodan locomotory mechanisms. Part 7. Functional requirements and body design in Colobognatha (Diplopoda), together with a comparative account of diplopod burrowing techniques, trunk musculature and segmentation. J. Linn. SOC. (Zool.), 44: , 35 text-figs., 3 pls. MANTON, S. M., Mandibular mechanisms and the evolution of arthropods. Phil. Trans. R. SOC. (B), 247: 1-183, 66 figs., 1 pl. MANTON, S. M., The evolution of arthropodan locomotory mechanisms. Part 8. Functional requirements and body design in Chilopoda, together with a comparative account of their skeleto-muscular systems and an Appendix on a comparison between burrowing forces of the arthropodan haemocoel. J. Linn. SOC. (Zool.), 45: , 99 text-figs., 7 pls. MANTON, S. M., The evolution of arthropodan locomotory mechanisms. Part 9. Functional requirements and body design in Symphyla and Pauropoda and the relationships between Myriapoda and Pterygota. J. Linn. SOC. (Zool.), 46: , 10 text-figs., 1 pl. MANTON, S. M., The polychaete Spinther and the origin of the Arthropoda. J. Nut. Hist.. 1: 1-22, 5 text-figs. MARTEN. W., Zur Kenntnis von Campodea. Z. Morph. bkol. Tiere, 36: 41-88, 29 text-figs. MATSUDA, R., Comparative morphology of the abdomen of a machilid and a rhidiid. Trans. Am. ent. SOC., 83: 39-63, 5 pls. MATSUDA, R., Morphology of the pleurosternal region of the pterothorax in insects. Ann. ent. SOC. Am.. 53: , 5 pls. MATSUDA, R., Some evolutionary aspects of the insect thorax. A. Rev. Ent.: 59-76, 3 text-figs. NOBLE-NESBITT, J., A site of water and ions exchange with the medium in Podura aquatica L. J. exp. Biol., 40: , 5 text-figs. OLROYD, H., The natural history of flies, xiv, 324, 32 pls., 46 figs. London: Weidenfeld & Nicolson. PARKER, T. J. & RICH, J. G., Observations on the myology of PalinumsEdwardrii Hutton. Proc. Linn. SOC. N.S. W. (Macleay Memorial Vol.): , 5 pls. PERRYMAN, J. C., The functional morphology of the skeleto-muscular system of the larval and adult stages of the copepod Calanus, together with an account of the changes undergone by this system during larval development. Thesis, University of London, 1-83, 28 figs. PRELL, H., Das Chitinskelett von Eosentomon. ein Beitrag zur Morphologie des Inseckten Korpers. Zoologica, Stuttg., Heft 64 (=Bd 25 Lfg 4), 1-58, 6 pls. ROEDER, K. D., The control of tonus and locomotor activity in the praying mantis (Mantis religiosa L.). J. exp. Zool.. 76: , 4 text-figs. ROLFE, W. D. 1. & INGHAM, J. K., Limb structure and diet of the Carboniferous centipede Arthropleura. Scott. J. Geol., 3: , 2 text-figs., 1 pl.

183 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 385 SCOURFIELD, D. J., The oldest known fossil insect (Rhyniella praecursor Hirst and Maulik). Proc. Linn. SOC. Lond., 152: , 10 text-figs. SEDGWICK, A., A student> text-book of zoology, i-xi, 1-905, 570 text-figs. London: George Allen. SHAROV, A. G., Peculiar Palaeozoic wingless insects of the new order Monura (Insecta Apterygota). Dokl. Akad. Nauk. SSSR, 122: SNODGRASS, R. E., Morphology and mechanism of the insect thorax. Smithson. misc. Coil, 80 (1): 1-108, 44 text-figs. SNODGRASS, R. E., The thoracic mechanism of a grasshopper, and its antecedents. Smithson. misc. Collns, 82 (2): 1-111, 54 text-figs. SNODGRASS, R. E., Morphology of the insect abdomen. Smithson. misc. Collns, 85 (6): 1-128, 46 text-figs. SNODGRASS, R. E A textbook of arthropod anatomy, 1-363, 88 text-figs. SNODGRASS, R. E., The evolution of arthropod mechanisms. Smithson. rnisc. Collns, 138: 1-77, 23 text-figs. SNODGRASS, R. E., Some words and their ways in Entomology. Proc. ent. SOC. Wash., 62: STURM, H., Die Nahrung der Proturen. Naturwhenschaften, 46: 90-91, 1 text-fig. TIEGS, 0. W., The development and affinities of the Pauropoda, based on a study of Pauropus sylvaticus. Q. JI microsc. Sci., 88: , 29 text-figs., 11 pls. TIEGS, 0. W., The flight muscles of insects-their anatomy and histology; with some observations on the structure of striated muscle in general. Phil. 7'runs. R. SOC. /B), 238: , 17 text-figs., 32 pls. TIEGS, 0. W. & MANTON, S. M., The evolution of the Arthropoda. Eiol. Rev., 33; , 18 text-figs. TILLYARD, R. J., The evolution of the Class Insecta. Papers Royal SOC. Tasmania, 1930: 1-89, 19 text-figs. TUXEN, S. L., The systematic position of entognathous Apterygotes. An. Esc. nac. Cienc. biol. Mix., 17 (1968): VERLUYS, J. & DEMOLL, R., Die Verwandschaft der Merostomata mit den Arachnida und die anderen Abteilungen der Arthropoden. Proc. Kned. Akad. Wet., 23 (5): , 6 text-figs. WEBER, H., Die Gliederung der Sternopleuralregion des Lepidopteren thorax. Eine vergleichend rnorphologische Studie zur Subcoxaltheorie. 2. wiss : , 21 text-figs. WEBER, H., Morphologie, Histologie und Entwicklungs-geschischte der Articulaten. Fortschr. Zool., 9 /1945-SO), , 24 text-figs. Appendix I THE MUSCULAR SYSTEM OF THE TRUNK OF THE JUMPING COLLEMBOLAN TOMOCER US LONGICORNIS The trunk musculature has been referred to above in general terms, sufficient to give an indication of the functions of the whole. But it is the detail of the musculature which shows how great are the modifications of the whole system in serving the needs of the springing organ. The differences between the trunk musculature of Collembola and of other apterygote classes is great, and functional needs account for these differences. Since the details are many, they are, for convenience, considered here, set apart from the main argument, but it is emphasized that on the details rests the general interpretation of the whole musculature of the apterygote classes. At the end of the descriptions of each category of trunk muscles given below is a summary of the functional significance of the details. The general conclusions concerning collembolan trunk muscles are recorded in 3 9B.
184 386 S. M. MANTON (i) The dorsal longitudinal system The plan of these muscles is shown diagrammatically in figure 19. The short muscles running from one intersegment to the next are shown on the left. They are all small, they lie in a more median (inner) series marked SI followed by the number of segment in which they lie, and an outer (more lateral) series, marked SO, also followed by the segment number. The muscles are labelled on figures 22A, B, 23D, 24G, H, 251, J and are unlabelled on other figures. The outer series extends only through the meso- and metathoracic segments and the first and second abdominal segments. The inner series extends further forwards and backwards. Muscle SI ThI-2 (figures 18a, 19 etc.) is the only sector of the dorsal system which passes through the prothorax and it does so without joining the immobile tendinous junction e. Muscle SI3 originates from a tendon arising from the dorsal cuticle just behind the abdominal 2-3 intersegment (figure 26N) and passes posteriorly (figures 26,O and 27P, Q) to end on the connective tissue covering the large muscle OE. Muscle S14 arises from the dorsal cuticle well behind the intersegment of abdominal segments, 3-4 (figure 27S), and ends on the following intersegment. These small rod-like muscles stain differently from the rest of the dorsal system, and probably are concerned with the maintenance of alignment between the segmental tergo-pleural arches. A series of long muscles passing through two consecutive segments is shown diagrammatically on the right of figure 19. These are bulky muscles which lie over one another except at their origins and insertions at the intersegments. They are shown typically on the first two abdominal segments and, as with the short muscles, form an inner (more median) and an outer (more lateral) series, marked I and 0, followed by letters in series from before backwards. Muscles IC and UC arise at the metathoracic-first abdominal intersegment and insert at the abdominal 2-3 intersegment. Muscles IB and OB from the metathorax insert at the abdominal 1-2 intersegment and muscles ID and OD from abdominal 1-2 intersegment insert at abdominal 3-4 intersegment. Thus each segment is traversed by 4 muscles which lie beside and over one another as shown in the sections, figures There is no mobile pro-mesothoracic intersegment, and muscles IA and OA, which would be expected to arise from such a junction if it was mobile, in fact arise from the meso-metathoracic intersegment and traverse the latter in the typical manner although the muscles are in fact short. Abdominal segments 3-5 show modifications connected with the jumping mechanism. Muscles ZE and OE from abdominal 2-3 intersegment arise typically and are very large. But, except for a large sector of muscle IE, they insert at the following intersegment without traversing a second segment. At the abdominal intersegment 3-4 muscles IE and OE are situated median to muscles ID and OD. Thus four large muscles instead of two insert on this intersegment and must exert considerable tension of tergo-pleural arch 4. In the fourth abdominal segment the corresponding large muscle F is undivided into inner and outer sectors and is short, inserting on the following intersegment 4-5. In the fifth abdominal segment these muscles are also undivided and short, marked IOG on figure 19, and G on figure 27T and U. Thus the most noteworthy features of the dorsal system are: the series of long muscles extending across two intersegments; these muscles are not
185 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 387 attached to the face of the tergo-pleural arches (see p. 307) but to their anterior margins or to the tendon system connected therewith; and the double layer in which these muscles lie, so promoting a maximum force which can shorten the body slightly. The small, rod-like, short muscles maintain exact alignment between the segments. Particular assistance to the jumping mechanism is provided by the length of the third abdominal segment and its muscles, together with the arrangement of muscles ID, OD, IE and OE which can effect a small but very strong displacement of tergo-pleural arch 5, thus shortening the body and increasing the internal hydrostatic pressure near the springing organ. The more posterior stout muscles F and IOG probably maintain stability and prevent intersegmental bulging under increase of hydrostatic pressure. The absence of any substantial dorsal longitudinal muscles in the pro- and mesothorax is due to the absence of mobility between the fused tergo-pleural arches of these segments and to the anterior attenuation of general shape so that the lateral longitudinal muscles maintain the stability of the metathoracic tergo-pleural arch, leaving unimpeded access of the tergal face for the extrinsic leg muscles of the first leg, which, in this narrow anterior end of the body, lie internal to the lateral and dorsal longitudinal muscles and not external to them, as on the meso- and metathorax (figure 18a pr.co.t. and ret.cot.t. from leg 1, etc. figures 22A, B, cf. figures 23D, F, 24G, 251). (iil The lateral longitudinal system These muscles are shown diagrammatically in figure 20b and typically consist of short muscles traversing one segment only. There are 5 distinct sectors on most segments, marked 1-5. Sector 5 in the metathorax and in abdominal segments 1-3 is small and rod-like, resembling the dorsal muscles of the short series (SO, SI etc.> and probably also maintains alignment (figures 25J, K, 24G, H, 26, 27P). In the third abdominal segment the posterior end of sector 5 is borne by a small tendon passing to the intersegment along with the deep oblique muscle ob of this segment. The other sectors are substantial, marked by numbers 1-4 in the sections on figures Anteriorly the whole series is attached to tendon e, representing the immobile pro-mesothoracic junction, and in front of this the lateral longitudinal muscles converge to insert on the intucked anterior margin of the prothoracic tergo-pleural arch to each side of the middle line, as indicated in figures 18a and 22A, both showing the muscle obliquely. Sector 1 of the lateral system is of particular interest. It is the most ventral of the series and is very large in the third abdominal segment from which it passes through the fourth abdominal segment to insert on the antero-lateral margin of the manubrium of the springing organ, forming the principal flexor muscle which folds up the organ after a jump (figure 17a). The origin of muscle I is shown superficially in figure 26M and its great size in figures 26N, 0, 27P, S and its insertion, ringed, in T. In the second and third abdominal segments sectors 2 and 3 form an inner series, attached to the deep tendon system at the intersegments (see 8 9A(v) and figures 26L, M, N, 0, 27P). In the third abdominal segment sector 4 is particularly large and spreads dorsally over part of the dorsal longitudinal
186 388 S. M. MANTON system to insert at the 3-4 intersegment beside the huge deep oblique muscle ob of segment 3 (figures 20b, 21a, 27P). In abdominal segment 4 the series 2-5 cannot be recognised and is represented by two large muscles marked 6 and 7 on figure 20b. The outer muscle 6 leaves abdominal 3-4 intersegment by a wide origin and converges on tendon q at the junction of the postero-lateral margin of the manubrium and segment 5 below its tergo-pleural arch (figures 20b, 17a, 27s). The inner muscle 7 arises internal to, as well as more dorsally than sector 6, and passes directly backwards to end on the antero-lateral margin of the 6th abdominal segment (figures 20b, 27S, T, U). In abdominal segment 5 only one pair of small lateral muscles are present additional to muscle 7. A small muscle v arises from tonofibrils near the origin of tendon dv in this segment and passes backwards to end on the lateral margin of the 6th abdominal segment (figures 20b, 21a, 27T, U). Particularly noteworthy features of the lateral longitudinal system are : the serial repetition of sector 5 promoting alignment between the tergo-pleural arches, and the position of the other sectors, which complete laterally the muscular cylinder whose contraction can increase the general hydrostatic pressure; the great size of sector 4, and the position of deep sectors 2 and 3 in the third abdominal segment, which can fortify the dorsal specializations leading to increase of hydrostatic pressure near the springing-organ ; the specialization of sector 1 in the third and fourth abdominal segments to form the flexor of the springing-organ; the modifications in the hinder abdominal segments which provide an anti-inflation arrangement which will maintain stability and prevent distension of the parts on increase in hydrostatic pressure. Any small pulling inwards of tendon q will contribute to the thrust exerted by the spring, but as the mechanical advantage is almost nil this must be a minor effect. (iii) The sternal longitudinal system These muscles are complex and composed of short and long sectors, and unlike the dorsal and lateral systems, they pass forward to the head, inserting on the head endoskeleton (as shown by Manton, 1964, figures 34, 35), traversing the flexible neck between head and prothorax, and marked A on figure 21a here. The short series of muscles are marked A-E on figure 21a and on the sections in figures They run between the thoracic endoskeletal plates g, backwards to the ribbon tendinous junctions. Sector D passes from the posterior part of the metathoracic plate g (figure 25I), through the first abdominal segment, (figure 255, K), to end on the ribbon tendinous junction in the anterior part of the second abdominal segment. Sector E arises from this junction and passes backwards to the ribbon junction of the deep dorso-ventral muscle dv in the anterior part of the third abdominal segment (figure 26N). SectorF leaves this junction and passes backwards to the distal end of the hamula (figures 21a, 26N, 0, 27P), and is probably homologous with the preceding short sectors. There are no more posterior short sectors, because of the presence of the basal plate and specializations connected with the springing-organ.
187 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 389 A plan of the long sternal muscles traversing more than one segment is shown in figure 21b, drawn with as little overlap as possible. The real postions of the sectors is clear in the sections. The whole system is arranged so that a maximum pull can be exerted by sectors H and J on the antero-ventral corner of the tergo-pleural arch of the fourth abdominal segment and by sector Gb on the margin of the basal plate, the ends of sectors H and J (figures 25K, 26, 27P) being in the cuticular furrow at the site named above. Sector Gb ends as shown in figure 27R between processes of tendon n just lateral to the basal plate. Sector Ga inserts close to tendon m just in front of the hamula (figure 26,O). Tendons m and n both spread the tergo-pleural arch tension to the ventral furrow without permitting a bulging of the ventro-lateral folds of the body wall. Sector H arises from endoskeletal plate g in the metathorax and opposite, on the other side of the plate, inserts sector X from the pro-thoracic plate and from this the sectors A extend direct to the head endoskeleton. Muscle Y from the prothoracic endoskeletal plate similarly supports the mesothoracic plate opposite to the origin of sector G which divides posteriorly into Ga and Gb. Long sector J arises from the ribbon tendinous junction bearing deep dorso-ventral muscle dv at the anterior end of the second abdominal segment, and close beside this junction, spreading into the posterior part of.the first abdominal segment is another tendinous ribbon junction bearing muscles Ka and Kb as shown in figures 21b, 255, K. Sector Ka arises from the posterior end of the mesothoracic endoskeletal plate (figure 24G by the upper g) and sector Kb passes forwards to insert on the stout endoskeletal strut K of the same segment (figure 23E), the muscle ending a little in front of the level of this section. There are no sternal longitudinal muscles in the abdominal segments 4-6 owing to the presence and movements of the springing organ. Thus the outstanding features of the sternal longitudinal muscles are: the long sectors, which exert particular tension at the posterior ends of muscles Ga, Gb, H and J (figure 21b), leaving the whole ventral region beyond these points free and under the influence of (i) hydrostatic pressures generated by the whole of the longitudinal and deep oblique systems, and (ii) the muscles concerned with stability and recovery movements of the springing organ. The short sectors of the sternal longitudinal muscles and the endoskeleton maintain the necessary stability for tension to be exerted by the long sectors. (iv) The deep dorso-ventral and deep oblique muscles These are very well developed and very simple. They arise as shown in figure 21a, one of each arises from the posterior end of the prothoracic endoskeletal plate, figures 18a, 22C (muscle dv sometimes in several sectors), and insert respectively on the dorso-lateral mesothoracic part of the common tergo-pleural arch (posterior to the extrinsic muscles of leg 1 and posterior to tendon e representing the thoracic 1-2 intersegment (figures 18a, 220, and on to the anterior margin of the metathoracic tergo-pleural arch. The series continues along the body arising from the tendons already described ( 8 9A(v) and figures 24G, 25J, 26L, N); muscles dv and ob are very large indeed in the long third abdominal segment. In the fourth abdominal segment only muscle dv is present (figures 21a, 27s) forming a very stout muscle pulling directly upwards from the sclerotized basal rod on the basal plate. In the fifth
188 390 S. M. MANTON abdominal segment muscles dv and ob appear to be represented by a tendon marked dv on figure 21a from tendon q and a fan of muscles s, t, u from the same site to the anterior edge of segment 6 (figure 17a). The absence of a deep oblique muscle ob in the fourth abdominal segment is due to the folding of the manubrium, as in figure 15b, the folding of the adjacent parts and the absence of a suitable site of origin for such a muscle. As has already been noted, contraction of muscle dv in segment 4 will assist the elasticity of the basal rod in reaching an extended position (figure 15a) from the curved resting position in figure 15b and so promote rapidity and force to the backward movement of the springing organ. Thus the great size, including length, of the deep oblique muscles in the third abdominal segment, suggests that they are of importance in augmenting a slight shortening of the body produced by contraction of the longitudinal muscular cylinder, both of which contribute to the sudden increase in hydrostatic pressure to the springing organ. In Part 8, p. 304 and figures 38, 39 the possible mode of action of the deep oblique muscles is considered in relation to the specializations shown by the dorsal system of Scolopendromorpha, the basic plan of chilopodan musculature being given on p No particular comments were made concerning the functions of the dorso-ventral muscles of Chilopoda besides noting that they must be antagonistic to the longitudinals and deep obliques and cause elongation of these muscles when relaxed after their contraction. The same is probably true for the jumping Collembola. The independent origins of the deep dorso-ventral and deep oblique muscles dv and ob of abdominal segment 3 in Tomocerus (figure 21a) is probably concerned with this antagonism. The oblique muscle arising anterior to the dorso-ventral muscle increases the length of the former, so facilitating a greater absolute shortening than would be possible to a shorter muscle, and the crossing of muscle dv and ob, muscle dv arising behind ob from the ventral cuticle, must facilitate the antagonistic action of the former. (v) The superficial pleural muscles These are shown diagrammatically in figure 20a. They all appear to be concerned with resisting any tendency to a bulging outwards by the body wall on increased hydrostatic pressure caused by contraction of the longitudinal and deep oblique systems. These muscles may be taken as typical in the first two abdominal segments. Two main sectors arise from the body wall just below the margin of the tergo-pleural arch and lateral to the ventral tube (figure 255, K, P1.1.) and fan upwards and forwards to the lateral face of this arch as shown. The tendon of origin of these muscles is inconspicuous in the first abdominal segment, but on the second it is in the form of a large ribbon tonofibrillar junction. On the third abdominal segment the pleural muscles are enormous, arising from the groove between the basal plate and tergo-pleural arch (figure 27P, R, P1.3.). One sector fans forwards to the anterior margin of the tergo-pleural arch (figure 26M, N, 0, P1.3.) and the other fans upwards as shown in figure 20a. They must augment the force exerted by sternal muscles Ga and Gb (figure 21b) in holding in the margin of the basal plate under increase of hydrostatic pressure. The pleural muscle of' the fourth abdominal segment forms a single stout
189 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 391 sheet arising from the vertical furrow between basal plates 4 and 5 (figure 15a, 20a, PZ.4.) and passes forwards and upwards to insert on the anterior end of tergo-pleural arch 4. Part of the origin of the muscle is cut in section figure 27S, and the muscle lies internal to the flexor muscle of the springing organ, marked 1. The pleural muscle of the fifth abdominal segment is in two sectors, as on the first to third abdominal segments. They arise from a thickened piece of highly sclerotized cuticle situated at the intucked edge of tergo-pleural arch 5 close to the posterior hinge of the manubrium (figures 15a, 20a, PZ.5.). The large sector is substantial and passes forwards through the fifth and fourth abdominal segments to end on the anterior margin of the tergo-pleural arch 4 as shown in figure 20a, the muscle is cut in section (figure 27S, P2.5.); the smaller sector passes dorsally to the face of the fourth abdominal tergo-pleural arch (figure 20a) and is shown, but unlabelled, inserting on the cuticle just dorsal to muscle PZ.5. in figure 27s. In the thorax two muscles appear to correspond with the series of pleural muscles present in the abdomen. They are inserted mainly upon the two thoracic tergo-pleural arches, and apparently represent the pleural muscles of the meso- and metathorax. The former consists of two sectors, as in most abdominal segments. One sector inserts on tendon e which represents the immobile anterior margin of the mesothoracic tergo-pleural arch, and the other inserts on the lateral face of the arch, as indicated in figures 18a, 20a. Both show typical insertions, and arise from a deep infolding of cuticle across the anterior proximal margin of the coxa of the second leg (figures 18a, 23D marked pleural mus. thorux2). It is a very stout muscle which must control any tendency of the flexible pleuron to bulge out between the first two pairs of legs, and of the second pair of legs themselves, on increase of hydrostatic pressure. A similar function must be exerted by the short very stout lateral longitudinal muscles of the prothorax passing from tendon e to the intucked anterior margin of the tergite (figures 18a, 20b, 22A). The maintenance of the intucking round the margin of the pro-mesothoracic tergo-pleural sclerite must prevent undue outbillowing of the flexible body wall below it. The interpretation of the pleural muscle of the metathorax is less certain. It has been shown that the pleural muscles of the abdomen arise on either side of the middle line on the ventral part of the body wall and the mesothoracic pleural muscles do so too. On figure 24H is shown a muscle marked pleural muxthorax 3 which arises opposite the middle of the third coxa, that is in the middle of the metathorax corresponding to the positions of the pleural muscles of the first, second, third and fifth abdominal segments. It arises from an intucking of the sternal cuticle at the proximal coxal margin where there is also situated a tendon of insertion of the long depressor from the trochanter dep. tr. I. The pleural muscle passes out to the lateral cuticle of the tergo-pleural arch, as on the mesothorax. Owing to the absence of a coxa-body articulation and the presence of endoskeletal struts p. and r, this pleural muscle can act as a strong coxal adductor muscle. In the mesothorax a separate coxal adductor muscle is quite differently contrived (figure 23D, Dd, E add). There is a small additional pair of pleural muscles in the metathorax, extending from the proximal anterior margin of the pleurite to the tergo-pleural arch above, so holding in the edge of the pleurite and preventing it
190 392 S. M. MANTON bulging out from the body (figure 18a, pleuralmus.). There are a number of small pleural muscles which control outbulging of all the flexible parts of the cuticle, for example, some sectors run with the coxal protractor of the first leg and arise on different parts of the cuticle above and in front of the coxa of this leg (figure 18a). Here there is a considerable expanse of flexible cuticle, well held by ties of various kinds (see Appendix 11), and bulging slightly in between them, figure 22A, ab marking the coxa of leg 1, the extend of the coxa is indicated by the tracing on figure 15d. Thus the major series of superficial pleural muscles shows remarkable serial uniformity and large size. They perform the same function everywhere, that of holding in the flexible ventral parts of the body wall against increase of hydrostatic pressure, and in this they are assisted by tendons rn and n in the abdomen which bear no muscles. Tendons m and n arise on either side of the mid-ventral line, anterior and posterior to the hamula, and insert on the ventral part of the tergo-pleural arch; tendon n passes just behind the shorter sector of pleural muscle 3 (figures 20a, 26,0, 27R). The ventral end of tendon n arises from the anterior extent of the cuticular basal rod in the basal plate (figure 15c). These tendons must firmly hold the ventro-lateral ridges of the body wall on either side of the ventral groove (shown in pl. 4, figure a, the ridges on either side of g), thus strengthening the actions of the pleural muscles in the vicinity of the hamula where very precise movements are needed (see below). The site of origin of the fourth pleural muscle in the abdomen is clearly associated with the ability to fold the springing organ at the end of a jump. If the sclerite labelled basal plate 5 on figure 15a really appertains to segment 5, then the position of origin of pleural muscle 4 is slightly atypical but functionally essential. If this sclerite really belongs to segment 4, then pleural muscle 4 is in the normal morphological position. The whole series of pleural muscles is associated with maintenance of body shape under increase of hydrostatic pressure and therefore assists in directing this force towards working the springing organ. (vi) The muscles of the hamula The median hamula is usually considered to represent the limbs of the third abdominal segment. The organ impinges in the notch at the base of the dentes of the springing-organ when it is folded forwards (figure 17V, unlabelled), so resting the flexor muscles and holding the organ against the elasticity of the cuticle of the bent basal rods of the basal plate (figure 15b, and see 0 9A(i)). Very delicate but small movements of the organ must take place, and the musculature is suited to this end. Distension of the organ giving increased projection from the body must be caused by hydrostatic pressure, implemented by trunk musculature. Retraction is mediated by the muscles F, the most posterior pair of short sectors of the sternal longitudinal muscles, shown in figures 21a, 26N, 0, 27P, inserting at the tip of the hamula. A complex of muscles, supported by tendinous junctions, controls the exact position of the hamula, set as it is in flexible sternal cuticle. Two pairs of tonofibrillar junctions with the sternal cuticle lie at the base of the organ and
 bear a series of muscles, both of which coverge to another tendinous junction (unlabelled) situated ventral to muscle ob on either side.")
191 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 393 more laterally (figure 27P). Short tendons from each (marked on the figure) bear a series of muscles, both of which coverge to another tendinous junction (unlabelled) situated ventral to muscle ob on either side. From this junction a long narrow muscle passes upwards and posteriorly to insert on the anterior margin of the tergo-pleural arch 4 (figure 17a). Another sheet of muscles pass from the last named tendinous junction downwards to the ventral body wall behind tendon n (figure 27R). The muscles of the hamula, with the exception of muscle F, cannot be related to any of the serially repeated components of the trunk musculature. They may represent the metamorphosed extrinsic muscles of an original pair of limbs. (vii) The muscles of the ventral tube The ventral tube is used for absorption of water and ions (Noble-Nesbitt, 1963). Distally the tube is provided with suctorial cups lined by extremely large unicellular gland cells and thin cuticle (figure 25J-K). Along the anterior face of the organ from the tip to its base and then along the mid-ventral line of the thorax, a gutter is formed in the cuticle, the side walls consisting of flanges staining bright red with Mallory and shown here in black (figures 16,1, 22-25,I). The gutter ends at the median gap in the labrum (Manton, 1964, figure 35) where presumably a secretion from the large salivary glands empties. Since the ventral tube is present in Collembola which have no springing organ, it cannot be concerned with the jumping mechanism. The function of the gutter is unknown. The ventral tube can be hydrostatically inflated (see figure 25J-K). The abundant longitudinal muscles of this organ prevent any undue dilatation during increase of hydrostatic pressure at a jump, and can retract the tube to a small lump on the ventral side of the body. The muscles are in three series. (a) Paired anterior and posterior median muscles arising from the middle of the main distal cup, the anterior muscle ending on a tendinous junction, supported by a cone of tonofibrils from the cuticle beside the ventral tube (figure 255). This same tendinous junction bears another group of tonofibrils (labelled) which unite with the strut bm from the end of the endoskeletal plate g of the metathorax. The posterior median muscle of the ventral tube passes upwards and laterally to reach the pleuron between the insertions of the two sectors of the pleural muscle, figure 25K, P1.I. (b) Three pairs of inner muscles arise from the lateral cups; they are cut short in figure 25K and entire in J. Muscles 1 and 2 insert above on to the hinder extension of the metathoracic endoskeletal plate g and muscle 3 inserts on the tendinous junction bearing the anterior median muscle of the ventral tube (figure 25J). (c) Intrinsic muscles are lateral in position. The proximal and lateral muscles are marked and inserted as shown in figure 255 and K, and may be concerned with alterations in direction of the projection of the ventral tube. It is noteworthy that all the extrinsic muscles of the ventral tube have very firm insertions outside the organ, suggesting a good ability to control unwanted inflation when the hydrostatic pressure increases during a jump. If the ventral tube represents the limbs of abdominal segment 1, then the extrinsic ventral
 Suspensory muscles from the coxa to the endoskeleton From the coxa of leg 1 two suspensory muscles sus.l. and sus.2.")
192 3 94 s. M. MANTON tube muscles would appear to represent metamorphosed extrinsic limb muscles because they do not correspond with parts of the basic trunk musculature. (viii) Suspensory muscles from the coxa to the endoskeleton From the coxa of leg 1 two suspensory muscles sus.l. and sus.2. pass from the lateral face of the coxa to the node G and to the plate g behind it, as shown in figure 18a and foreshortened in figure 22B. The muscles lie at an angle to one another. From the distal coxal segment b of leg 2 three suspensory muscles sus. 2., sus. 3. and sus. 4. leave the lateral face and insert on and around the node G (figure 18a and 23E). Muscle sus.1. passes upwards and forwards to the posterior end of endoskeletal plate g of the prothorax (figures 18a, 23D, E). Muscles sus. 2, sus. 3. and sus. 4. are similarly present on leg 3 but there is here no sus. 1. muscle. The presence of the latter on leg 2 is probably correlated with the leg-movements taking place both forward and backward of this coxa-body joint. Appendix I1 THE CUTICULAR CONNECTIONS OF THE THORACIC ENDOSKELETAL SYSTEM OF TOMOCER US LONGICORNIS There is a tendency among some entomologists to regard the endoskeletal structures existing among the hexapod classes as being homologous with one another (Carpentier, 1949, etc.), regardless of the component parts being formed by (i) apodemes, (ii) connective tissue endoskeleton or (iii) muscles. These views are not supported by the present work (9 9B, 10D). Similarly, the many ties between the connective tissue endoskeleton and the cuticle are presumed by some to be homologous from class to class, and the resemblances are regarded as indicative of phylogenetic affinity. In the present account the differences between the endoskeletal structures of the several hexapod classes have been stressed, and it has been suggested that resemblances between endoskeletal structures of different composition may be explicable on a supposed convergent meeting of common functional needs. The ties, or stabilising struts, which connect the thoracic endoskeleton with the cuticle, are various and among the apterygote classes they are most elaborate and numerous in the Collembola where the need to secure anchorage for the tendinous endoskeletal plates is very great, particularly at the moment of increased muscular tension at a jump when the long sternal longitudinal and deep oblique muscles contrast (0 9A(iv), (v)). The collembolan endoskeletal structures are anchored not only by the presumably persistent intersegmental ties remaining after the embryonic formation of the endoskeleton, but by ties ranging from compact cones, or short ribbons, of tonofibrils converging on to endoskeletal plates, to fibrils concentrating into long struts, and presumably developed from the general subcutaneous layer of connective tissue as well as by tonofibrils. It is shown below how different is the complement of ties on the prothorax of Tomocerus from those on the meso- and metathorax. Since
193 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 395 such differences can exist in consecutive segments of the same animal it is doubtful whether phylogeny can account for resemblances between particular segments of species from different classes. Such resemblances are more likely to be due to similar functional needs rather than to the persistence of any basic type of organization. The large dorsal and lateral ties between the thoracic endoskeleton of Tomocerus have been described above (6 9A(v)). The numerous ventral ties are recorded below. Elucidation of the nature of these ties needs Azan or Mallory s triple staining of serial sections in order to differentiate decisively between brown-staining unstriated tonofibrils contained within ectodermal cells, which are often enormously enlarged (see ventral end of tendon rn, figure 26,O) and true purple-staining muscle fibrils which are striated and not contained within ectodermal cells. The connective bm on the meso-metathorax junction of Tomocerus Zongicornis (figure 23F) is not entierement musculaire as is claimed by Carpentier (1949) for the similarly placed tie in the collembolan Tetradontophora gigas. It is probable that tonofibrillar bundles of large size have been considered to be muscles by Carpentier because his technique did not distinguish between the two. Similarly the long tie q in Tomocerus longicornis (figures 23D, 24G) is tonofibrillar and not muscular, as claimed for the similarly placed tie in Tomocerus plumbeus and Tetradontophora gigas by Carpentier. In order to determine how much, if any, phylogenetic significance can be attached to the details of endoskeletal connection, and for accurate comparison to be made with comparable structures in other classes, the endoskeletal ties between the endoskeleton and cuticle of the entire thorax of Tornocerus Zongicornis are listed below. The abdominal connections are recorded in 5 9A(v). Thoracic structures are shown by thick sections in figures viewed from in front, but the slopes and exact levels of some of the ties and muscles cannot also be shown (see Carpentier, 1949). In Tomocerus the prothoracic endoskeletal plates are fused mid-ventrally where the legs are close together (figure 22A); in Tetradontophora Carpentier has shown that this median continuity exists here on the meso- and metathorax also. On the meso- and metathorax, using the labelling of Carpentier (1949), the most anterior of the ventral ties between the paired endoskeletal plates and the cuticle is tie q, from the anterior end of each plate and reaching to the middle line, figures 23D, 24G. Behind it lies tie f to the side of the ventral gutter, taking some of the strain from the adductor muscles of the mesothoracic coxa (figure 23Dd). On the methathoracic segment, where there is no corresponding coxal adductor muscle, tie f arises from the sides of a median cuticular depression (figure 24Gg). Tie e lies entirely (mesothorax) or mainly (metathorax) lateral to the ventral gutter and opposite to the middle part of the coxa (figures 23, 24). Tie bm passes from the plate g dorsal to the nerve cord to insert on the cuticle at the apex of a median tubular depression at the thorax 2-3 intersegment (figure 23F). The posterior end of plate g on the metathorax is linked by a tie labelled bm, figures 25J, which probably corresponds with that of the mesothorax. It joins the cuticle lateral to the mid-ventral line behind coxa 3 and near the origin of the ventral tube. Another tie, s passes from the plate g in the mesothorax only to the proximal posterior margin of the proximal coxal segment (cut in two parts in figures 23E, F). On
 with the exception noted in 0 9A(v) and the additional three items just described.")
194 396 S. M. MANTON the metathorax a small rod leaves the anterior margin of the proximal coxal segment a to end on the connective tissue around the sternal muscles (figure 24G unlabelled). These findings correspond with the ties recorded by Carpentier (1949) with the exception noted in 0 9A(v) and the additional three items just described. There are other important processes from the plate g and its anterior node G which support limb muscles such as that to muscle sus.co.pz. shown in figure 23E unlabelled. The first pair of legs differs from the others in being set close together ventrally on the narrowing prothorax. A modified dorso-lateral prothoracic tie k is present, but does not reach the cuticle (figure 22B) and instead carries a muscle to the tergite. No lateral ties, such as r and p, are present more posteriorly. Tie r is associated with the mobility of the proximal coxal subdivision, complete or otherwise on legs 2 and 3, and is not required on leg 1 where the coxa is undivided. Tie p is also not required on the prothorax, there being no pleurite. Instead there are three antero-lateral ties marked I, 2, 3 to the pleuron (stippled in figure 22A) which bays out between them. Tie I passes to the dorsal connective tissue over the sternal longitudinal muscles and does not directly join the plate g. Ties 2 and 3 pass to plate g and to the endoskeletal node G respectively. The endoskeletal plates g are fused mid-ventrally at their anterior ends and linked here by a pad of tonofibrils, tie 4, to the cuticle in the mid-ventral line (figure 22A). This tie probably corresponds with paired ties q elsewhere. The main mass of the ventral longitudinal muscles passing forward to the head lie ventral to the main part of the tergo-pleural arch of the prothoracic segment, and it is thus not surprising to find the plate g linked with the pleuron and not with the tergo-pleural arch. Tie 5 passes from the sternal cuticle to the node G. Tie 6 may represent tie f of other segments (figure 22B cf. 23Dd, 24Gg). Tie 7, with tie 8 a short distance posterior to it (figure 22Bb, C), seem a little too posterior to represent tie e of other segments (figures 23E and 24H). Tie 9 is lateral in position and just posterior to the junction of coxa and body (figure 22C) and is not represented on other segments. Tie 10 (figure 22C) is posterior in level to tie 8 and passes dorsal to the nerve cord to unite with the apex of a mid-ventral intersegmental depression. Tie I0 is directly comparable with tie brn of other segments (figure 23F). Thus very few of the prothoracic ties can reasonably be equated with those on the meso- and metathorax. The prothoracic equipment in ties suits the needs of this segment, and it is highly probable that these endoskeletal ties to the cuticle are present just where they are required for mechanical reasons. Since the ties can be so different on two consecutive segments of the same animal, it is unwise to attach much phylogenetic significance to similarities in endoskeletal connections between thoracic segments of animals belonging to different classes. Such similarities are likely to have a functional rather than a phylogenetic explanation.

195 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANlSMS 397 EXPLANATION OF PLATES The black or white spots placed to the side of a limb tip indicate that this leg is in contact with the ground and is performing the propulsive backstroke. The black or white short horizontal lines placed near a limb tip indicate that the tip is off the ground and the leg is performing the forward swing. These marks are additional to the footprints forming the tracks left on smoked paper. The pattern of the gait, as in previous Parts, is expressed by the figures within the brackets in the legends, which give the relative durations of the forward and backward strokes, the duration of a pace being taken as 10. The phase difference between successive legs, p.d., is given as that proportion of a pace by which leg n + I is in advance of leg n, the duration of a pace being taken as 1.O. PLATE 1 (a-b) Forficula uuriculuriu L. 11 mm. (a) The earwig ran over smoked paper, leaving a track of footprints, and continued across white paper, lined at 2 cm distances. Gait (4.0: 6.0) approx., p.d. 0.45, the moment shown is marked on figure 2b by the arrowed line Pl.1~. All legs are in contact with the ground except left leg 3. The footfall of right leg 2 is lateral to and level with that of right leg 1, the tip of left leg 3 will be put on the ground just behind and between the levels of footfalls 1 and 2. Stride length 4.4 mm. (b) Track made by earwig running faster than in (a). Gait (5.0: 5.0) approx., p.d. 0.55, stride length 6.2 mm. The corresponding gait diagram is shown in figure 3a. (c) Shield bug Dolycoris baccarum (L.) 11 mm, walking over smoked paper, gait (4.0: 6.0) approx., p.d. 0.45; the moment shown is marked on figure 3c by the arrowed line PLIc. Left leg 2 and right leg 3 are off the ground. The individual footprints are most easily seen around left leg 3 and in the preceding set. Stride length 6.5 mm. (d-f) Cylindronotus laevioctostria%us Gaeze 8.8 mm. (d) Walking over smoked paper, gait (3.5 : 6.5) approx., p.d. 0.4, as in (e), where a similar moment is shown with left leg 3 only off the ground, stride length 4.4 mm approx. (f) Walking faster by gait (5.0 : 5.0) approx., p.d. 0.55, the moment shown is marked on figure 3a by the arrowed line Pl.If: Left leg 2 and right legs 1 and 3 are off the ground. (g-h) Staphylinid beetle larva 30 mm, digitigrade. (g) Walking over smoked paper. Successive groups of left footprints shown on the track are marked 1-5. Paired leg in roughly the same phase, footprints near thorax and near hinder abdomen are roughly level on the two sides, stride length here 11.2 mm, but can be half of this when abdominal punting is less vigorous. Behind the body there are three groups of footprints on the right in series with those alongside the body, on the left the groups are more forward in position so that they almost alternate in level with the right hand groups, indicating that the legs then were nearly in opposite phase. (h) Walking on white paper, paired legs in opposite phase, gait (2.0 : 8.0) approx., p.d. 0.33, the moment shown is marked on figure 2a by the arrowed line PLlh., only right leg 2 is off the ground. (i-k) Acerentomon nemorale Womersley 1.6 mm. (i, j) Walking on damp white paper, (k) walking on black paper. Gaits in all (2.0 : 8.0) approx., p.d. 0.25, the moments shown are marked on figure 2c by the arrowed lines Pl.1 k., i., j., right legs 3 in (i, j) are off the ground and left leg 2 in (k) is off the ground, the other three legs forming stable support in the quatrupedal gait in each case. (I-m) Petrobius brevisiyll Carpenter 12 mm, running on damp white paper, paired legs in similar phase, (I) Gait (6.7 : 3.3) approx., p.d. 0.33, legs 1 and 3 are off, and legs 2 on, the ground. (m) Gait (5.0 : 5.0) approx., p.d. 0.33, much as in pl. 5, figure a, probably with legs 1 on the ground, leg 2 rapidly swinging forwards and legs 3 just about to be put on the ground. The moments shown are marked on figure 8a, by the arrowed lines py.11.. P1.5~. respectively. (n) A sminthurid collembolan 1.7 mm, gait (3.0 : 7.0) approx., p.d PLATE 2 (as) Campodeu stuphylinus Westwood 4.5 mm, one antenna lacking its tip. (a) Running on damp paper by fastest recorded gait, (5.5 : 4.5), approx., p.d. 0.6, speed 54 mmlsec., stride length 3.0 mm approx., and pace duration 0.05 sec, determined by cinematography. The details of the gait are shown in figure 5a, c, e, the gait still 5g shows the legs on and off the ground, as in the photo. The fields of movement are marked by the vertical lines to the side, which do not overlap, as they do in reality, see fwre 5j. The moment in time is marked by the arrowed line and diagram on figure Sa, g. (b) Running on smoked paper by gait (3.5 : 6.5). p.d. 0.4 approx. giving speeds of < 26 mdsec as determined by cinematography, stride lengths 2.5 mm approx. and pace durations sec. Staggered sets of footprints are arranged as shown on figure lb. Each leg
196 398 S. M. MANTON oversteps the leg in front, the differential leg length bringing the footfalls of legs 2 and 3 outside and in front of those of the preceding leg. The gait still, figure 5h shows which legs are on and off the ground in the photo. (c) Running more slowly on smoked paper by gait (2.5 : 7.5), p.d approx., giving speeds of 18 mdsec or less, stride lengths 2.0 mm approx., pace durations variable and longer than in preceding gaits (a, b). Gait details are given in figure 5b, d the gait still 5i showing which legs are on and off the ground in the photo. There are slight irregularities in leg positions caused by reaction to sudden light intensity at the moment of photography. In (b) left leg 1 has slipped back from its footfall, forming a scratch as the head was turning to the left. Tracks with complete regularity of footprints are obtained in dim illumination. (d-i) A japygid sp. from the Solomon Islands, 9.2 mm (h-i) in less bright light and at a lower magnification than (dg). All show slow walking, gait (2.5 : 7.5) approx., p.d. 0.33, as in figure 5b, d,i, or with longer relative duration of backstroke. The antennae are not foreshortened, they are approximately parallel to the ground and their differences in apparent length are real, due to telescoping or extension of antenna1 segments; the shortest antenna, 2.4 mm is on the left of (d) and the longest 3.5 mm on the right of (e). (h) Shows a rare moment when this animal made a brief essay at fast walking using the same pattern of gait as Campodeu (a), the moment shown is a mirror image of (a). PLATE 3 (ad) Tomocerus longicornis Muller. 5 mm. (a) Running by its fastest gait (4.4 : 5.6). p.d. 0.44, the details of the gait are shown in figure 6a, d, g. Left leg 1 and right leg 3 are off the ground. (bd) Lateral views on a flat substratum and in (d) on a grass stem. (e) Walking over smoked paper by the same gait as in (a), the moment in time being slightly later, see arrow below figure 6a. (f) Orchesella villosa, Geof. 4 mm. Walking by the same gait as in (a, e), the moment in time being a little later than in (e) and is marked by the arrow below figure 6a. (g) Anuridu mantimu, (GuCrin) 2mm walking on damp ground glass by gait (3.3 : 6.7), p.d. 0.33, the moment in time is marked by the arrow below figure 6c and the gait still 6j shows which legs are on and off the ground in the photo. (h, i) Neanura muscorum Templeton 2 mm displaying the flexibility of body which is absent in the rigid-bodied, jumping Collembola (a-f). PLATE 4 Scanning electron micrographs of freezedried Collembola. (a-c, e, f) Tomocerus longicomis Muller, (d) OrckeseUn vium Geof. (a) Slightly oblique lateral view to show the ventral groove g along the abdominal segments into which the manubrium of the springing organ fits when impinged upon the hamula. x24. (b) Lateral view of another specimen of Tomocerus, tilted slightly in a ventro-lateral direction, ~40. A diagram of this micrograph is given in figure 15d. The thoracic and abdominal tergo-pleural arches are marked by numbers, as are the left thoracic legs. The anterior margin of the coxa ub of left leg 1 lies at the edge of the letter a; two folds of pleuron are situated to the left of this. The coxa is coloured quite differently from the pleurosternal folds of cuticle owing to the degree of sclerotization, thus the limits of the coxa are quite clear in sections and under the light microscope, but this distinction is not apparent on the electron micrograph. The margin of the tergo-pleural arch above leg 2 is masked by scales. The proximal coxal segment of leg 2 is obliquely set and marked a. The longer distal segment is marked b. On leg 3 the junction between the proximal and distal coxal segments lies at the row of scales just below the a marking the proximal segment. The distal segment is marked b. The pleurite PI. carries a row of conspicuous scales, and further scales obscure the edges of the tergo-pleural arch. The limits of the expanse of arthrodial membrane ar. above the aochanter are shown in the diagram in figure 15d. (c) Lateral view of the posterior part of the abdomen of Tomocerus, slightly tilted in the ventro-lateral direction, x80. The 3rd, 4th and 5th tergo-pleural arches are marked by numbers. The basal plate BP4 is curved in to form the ventral groove g: this folding leaves a conspicuous ventro-lateral point p, seen also in (a). The paired, lateral, close articulations between the basal rods and the manubrium M lies to the right of the +, the points marked p and + which lie nearest to the bottom of the page appertain to the animal s right side. The flexible zone between basal plates 4 and 5, BP4. BPS is marked f. (d) Direct lateral view of Orchesella, marked as for (c) with the manubrium more fully extended. The inwardly directed junction between basal plate 4 BP4 and tergwpleural arch 4 above it is plainly seen as a line. The dorsal part of basal plate 5 BP5 bears long setae while the
197 EVOLUTION OF HEXAPOD LOCOMOTORY MECHANISMS 399 ventral part, which tucks inwards on forward flexure of the manubrium, is nearly bare. x40. (e) Tomocerus.longicomis to show the base of leg 3 in lateral view at a higher magnification than for the specimen in (b). The pleurite pl. lies under its row of scales, the partially divided coxal segments a and 1, are separate on this side of the leg and are marked in white, part of the trochanter t and first abdominal segment abdl are labelled, and ar in black marks the flexible cuticle between coxa and trochanter. x112. (f) Tomocerus longicomis. To show the surface scales and spines at the region marked by a black ring on (b). Some scales are lost, so exposing the bare cuticle and its thorax 3-abdomen 1 intersegmental fold. x896. PLATE 5 (a, b) Pehobius breuisrylis Carpenter. (a) Specimen 10 mm, running over smoked paper by the jumping gait, with legs of each pair in similar phase, the footprints on the two sides of the track being level. The moment in time is unstable, with only one pair of legs (legs 1) in contact with the ground; a double claw mark is seen at the tip of right leg 1. The long exposure time, 1/60 sec, gives a blurred image of legs 2 as they rapidly swing forward, leaving a single claw mark. showing on the right as a white spot immediately in front of the double claw mark of leg 1, and on the left by a white spot in front and lateral to the footmark left by leg 1. Legs 3 have vacated their footprints lateral to those of legs 1 and 2 and are nearing their forward position when they will be put on the ground lateral to the foorprints of legs 1 and 2 in front. Legs 3 are not blurred and must be moving more slowly forwards than legs 2. The gait is (5.0 : 5.0) approx., p.d and the moment in time shown by the photo is marked by the arrow below the gait diagram in figure 8a. (b) Track of a specimen 6 mm, running by a faster gait, (6.0 : 4.0)-(6.7 : 3.3) approx., p.d. 0.33, in which the footfalls are staggered, cf. (a), and the stride (distance between the footprints on one side) is relatively longer, see figure 8h. The heavy white vertical lines on (a, b) record the body length of each animal, so that ready comparison may be made of the stride lengths relative to the body length. (c, d) Thermobia domesrica Pack, 5 mm, running over smoked paper, exposure time 1/500 sec. Paired legs are in opposite phase and the groups of footprints on opposite sides of the track are staggered and not level as in (a, b). (c) The heavy white vertical line records the body length, two stride lengths exceed it, cf. Pehobius running fast (b) and not so fast (a). The gait pattern is (6.7 : 3.3) approx., p.d The left and right legs correspond with the moment in time marked by vertical arrows L and R on figure 8a for gait (6.7 : 3.3). Left leg 1 is on the ground but invisible, left leg 2 is moving forwards off the ground and left leg 3 is about to be put down in front and outside the footprint of leg 2; right legs 1 and 3 are off the ground and right leg 2 is propulsive. The moment is unstable, with only two legs in contact with the ground, as in (a), but with paired legs in opposite instead of the same phase relationship. (d) Shows a moment in time a little later than in (c). Left leg 3 is on the ground, near the end of the backstroke, right leg 3 is about to be put down in front of the footprint of leg 2 when leg 2 swings forward, left leg 2 is about to be put on the ground, left leg 1 is invisible but probably moving forwards while right leg 1 is on the ground, there is irregularity in leg positions caused by sudden bright illumination at the moment of exposure, but the track is regular. (e-0 Hypogastrurid collembolan, (e) in dorsal view walking over smoked paper, (f) in ventral view seen through cover glass. The legs are short and 3% stride lengths are required to exceed the body length, cf. (a, b, c). (g-h) Petrobius brevisrylis Carpenter, successive frames of a cinematograph film in lateral view, the animal performing the jumping gait along a narrow ledge covered with damp filter paper marked by black lines 2 cm apart. The two horizontal lines in each photo show the limits of the ledge, the 2 cm distance being foreshortened. The vertical lines in the photos cross the ledge and rise over the vertical wall behind the ledge. The two frames show that one cycle of leg movements has been executed between the two exposures, the limb tip of leg 3, on the ground in each frame, is marked. Cinematograph film records of the animal running more slowly are given in pl. 6, figures b d taken under the same circumstances. PLATE 6 Thysanura, walking and jumping gaits. (ad) Frames from a cinematograph film in lateral view of Pehobius breuirtylis Carpenter 12 mm, progressing along a ledge, as described for pl. 5, figuresg, h. The paired legs are in similar phase. (a) The animal is stationary and resting. (bd) three successive frames of the animal coming to rest after a series of jumps along the ledge. In (a), the resting animal shows leg 3 in the extreme remotor position, leg 2 has been placed on the ground just anterior to the footfall of leg 1. The stride length is roughly the distance between footfalls 2 and 3, and is
 leg 2 has been placed on the ground, starting the backstroke, the tip being stationary; the thorax is thrown upwards and the abdomen downwards by the thrust exerted by legs 2.")
198 400 S. M. MANTON shorter than in (d), but the stride can be much longer than in either (a) or (d). In (b) leg 2 has been placed on the ground, starting the backstroke, the tip being stationary; the thorax is thrown upwards and the abdomen downwards by the thrust exerted by legs 2. Legs 1 and 3 are off the ground, but swinging forwards for their footfalls seen in (c, d). In (c) leg 2 is swinging forwards (blurred) and is almost upon the pound in (d); leg 3 is on the ground in (c, d), and its thrust exerted in (c) throws the head end down and the abdomen up; the movement is accompanied by dorsal concavity of the abdomen which must be effected by abdominal muscles. The head end advanced a little between frames (c) and (d) but there is no full remotor movement by the legs as the animal comes to a stop. (e-g) Petrobius brevirtylis running upside down on a damp filter paper ceiling. The paired legs are now in opposite phase, resulting in stability, with three or more legs in contact with the ground at all moments in time (figure 7f, g). The gaits are (4.0 : 6.0) approx., p.d in (e) and (g) and (2.5 : 7.5) p.d in (f), see figure 7b, c showing the gait diagrams of walking gaits. The moments in time of the photographs are marked by arrows below the gait diagrams in figure 7b, c. The gait stills showing the disposition of the legs on and off the ground in figure 7h, i, j, correspond with photos (e, g, f) respectively. (h) LepLma saccharina L. 8 mrn, running on smoked paper, m m slowly than Themzobia in pl. 5, figures c, d, and with more irregularity. The right and left groups of footprints are staggered in the lower part of the track, near to the animal they become almost level and at the moment of photography the movement of paired legs in opposite phase is restored. This demonstration of stronger unilateral than transverse coordination also occurs in Onychophora, Geophilomorpha, etc. Left leg 3 is just leaving the ground, right leg 3 is about to be put down, left leg 2 is on the ground while its fellow is not, left leg 1 is off the ground and right leg 1 is invisible, but probably propulsive. These positions correspond with a gait of about (5.0 : 5.0) p.d

199 S. M. MANTON (Facing p. 400)
 Plate 2 S. M.")
200 Zool. J. Linn. SOC., 51 (1972) Plate 2 S. M. MANTOIV
 Plate 3 s.")
201 Zoo/. 7. Linn. Soc.,.il (1972) Plate 3 s. n t MANTON
 Plate 4 S. M.")
202 Zool. J. Linn. Soc., 51 (1972) Plate 4 S. M. MANTON


203
 Plate 6 S. M.")
204 Zool, J. Lznn. Sac., 51 (1972) Plate 6 S. M. MANTON
Supplementary Figure 1 An insect model based on Drosophila melanogaster. (a)
") Supplementary Figure 1 An insect model based on Drosophila melanogaster. (a) Side and ventral images of adult female flies used to calculate the sizes of body and leg segments. Scale bar is 0.3 mm. Green,
Supplementary Figure 1 An insect model based on Drosophila melanogaster. (a) Side and ventral images of adult female flies used to calculate the sizes of body and leg segments. Scale bar is 0.3 mm. Green,
SOME ASPECTS OF THE MECHANICS OF MAMMALIAN LOCOMOTION
 SOME ASPECTS OF THE MECHANICS OF MAMMALIAN LOCOMOTION BY OLIVER R. BARCLAY Department of Zoology, University of Cambridge (Received 28 August 195a) Gray (1944) analysed the pattern of muscular activity
SOME ASPECTS OF THE MECHANICS OF MAMMALIAN LOCOMOTION BY OLIVER R. BARCLAY Department of Zoology, University of Cambridge (Received 28 August 195a) Gray (1944) analysed the pattern of muscular activity
CHAPTER IV FINITE ELEMENT ANALYSIS OF THE KNEE JOINT WITHOUT A MEDICAL IMPLANT
 39 CHAPTER IV FINITE ELEMENT ANALYSIS OF THE KNEE JOINT WITHOUT A MEDICAL IMPLANT 4.1 Modeling in Biomechanics The human body, apart of all its other functions is a mechanical mechanism and a structure,
39 CHAPTER IV FINITE ELEMENT ANALYSIS OF THE KNEE JOINT WITHOUT A MEDICAL IMPLANT 4.1 Modeling in Biomechanics The human body, apart of all its other functions is a mechanical mechanism and a structure,
Assessments SIMPLY GAIT. Posture and Gait. Observing Posture and Gait. Postural Assessment. Postural Assessment 6/28/2016
 Assessments 2 SIMPLY GAIT Understanding movement Evaluations of factors that help therapist form professional judgments Include health, palpatory, range of motion, postural, and gait assessments Assessments
Assessments 2 SIMPLY GAIT Understanding movement Evaluations of factors that help therapist form professional judgments Include health, palpatory, range of motion, postural, and gait assessments Assessments
-Elastic strain energy (duty factor decreases at higher speeds). Higher forces act on feet. More tendon stretch. More energy stored in tendon.
. Higher forces act on feet. More tendon stretch. More energy stored in tendon.") As velocity increases ( ) (i.e. increasing Froude number v 2 / gl) the component of the energy cost of transport associated with: -Internal kinetic energy (limbs accelerated to higher angular velocity).
As velocity increases ( ) (i.e. increasing Froude number v 2 / gl) the component of the energy cost of transport associated with: -Internal kinetic energy (limbs accelerated to higher angular velocity).
Complex movement patterns of a bipedal walk
 1 Complex movement patterns of a bipedal walk Objectives After completing this lesson, you will be able to: Describe the complex movement patterns of a bipedal walk. Describe the biomechanics of walking
1 Complex movement patterns of a bipedal walk Objectives After completing this lesson, you will be able to: Describe the complex movement patterns of a bipedal walk. Describe the biomechanics of walking
Arthropods are everywhere
 Phylum Arthropoda chitin exoskeleton jointed appendages repeated segments open circulatory system more than 10 million species respiratory system varies with subphlyum Arthropods are everywhere Two out
Phylum Arthropoda chitin exoskeleton jointed appendages repeated segments open circulatory system more than 10 million species respiratory system varies with subphlyum Arthropods are everywhere Two out
BIOMECHANICAL MOVEMENT
 SECTION PART 5 5 CHAPTER 12 13 CHAPTER 12: Biomechanical movement Practice questions - text book pages 169-172 1) For which of the following is the athlete s centre of mass most likely to lie outside of
SECTION PART 5 5 CHAPTER 12 13 CHAPTER 12: Biomechanical movement Practice questions - text book pages 169-172 1) For which of the following is the athlete s centre of mass most likely to lie outside of
THE OHIO JOURNAL OF SCIENCE
 THE OHIO JOURNAL OF SCIENCE Vol. 72 JULY, 1972 No. 4 BODY FORM AND GAIT IN TERRESTRIAL VERTEBRATES 1 WARREN F. WALKER, JR. Department of Biology, Oberlin College, Oberlin, Ohio 44074 One appeal of pure
THE OHIO JOURNAL OF SCIENCE Vol. 72 JULY, 1972 No. 4 BODY FORM AND GAIT IN TERRESTRIAL VERTEBRATES 1 WARREN F. WALKER, JR. Department of Biology, Oberlin College, Oberlin, Ohio 44074 One appeal of pure
STUDIES IN ANIMAL LOCOMOTION
 39 1 STUDIES IN ANIMAL LOCOMOTION III. THE PROPULSIVE MECHANISM OF THE WHITING (GADUS MERLANGUS) BY J. GRAY. (From the Laboratory of Experimental Zoology, Cambridge.) {Received ^th February, 1933.) (With
39 1 STUDIES IN ANIMAL LOCOMOTION III. THE PROPULSIVE MECHANISM OF THE WHITING (GADUS MERLANGUS) BY J. GRAY. (From the Laboratory of Experimental Zoology, Cambridge.) {Received ^th February, 1933.) (With
Coaching the Triple Jump Boo Schexnayder
 I. Understanding the Event A. The Run and Its Purpose B. Hip Undulation and the Phases C. Making the Connection II. III. IV. The Approach Run A. Phases B. Technical Features 1. Posture 2. Progressive Body
I. Understanding the Event A. The Run and Its Purpose B. Hip Undulation and the Phases C. Making the Connection II. III. IV. The Approach Run A. Phases B. Technical Features 1. Posture 2. Progressive Body
Walking and Running BACKGROUND REVIEW. Planar Pendulum. BIO-39 October 30, From Oct. 25, Equation of motion (for small θ) Solution is
 Solution is") Walking and Running BIO-39 October 30, 2018 BACKGROUND REVIEW From Oct. 25, 2018 Planar Pendulum Equation of motion (for small θ) 2 d g 0 2 dt Solution is Where: g is acceleration due to gravity l is pendulum
Walking and Running BIO-39 October 30, 2018 BACKGROUND REVIEW From Oct. 25, 2018 Planar Pendulum Equation of motion (for small θ) 2 d g 0 2 dt Solution is Where: g is acceleration due to gravity l is pendulum
A Hare-Lynx Simulation Model
 1 A Hare- Simulation Model What happens to the numbers of hares and lynx when the core of the system is like this? Hares O Balance? S H_Births Hares H_Fertility Area KillsPerHead Fertility Births Figure
1 A Hare- Simulation Model What happens to the numbers of hares and lynx when the core of the system is like this? Hares O Balance? S H_Births Hares H_Fertility Area KillsPerHead Fertility Births Figure
Motile Organisms. Name: Lab section: BACKGROUND READING: This lab handout Chapter 49, pp
 Name: Motile Organisms Lab section: BACKGROUND READING: This lab handout Chapter 49, pp. 1011-1014 PRE-LAB: Do the pre-lab reading and carefully read through the lab. Come to lab with a clear idea of what
Name: Motile Organisms Lab section: BACKGROUND READING: This lab handout Chapter 49, pp. 1011-1014 PRE-LAB: Do the pre-lab reading and carefully read through the lab. Come to lab with a clear idea of what
THERMALLING TECHNIQUES. Preface
 DRAFT THERMALLING TECHNIQUES Preface The following thermalling techniques document is provided to assist Instructors, Coaches and Students as a training aid in the development of good soaring skills. Instructors
DRAFT THERMALLING TECHNIQUES Preface The following thermalling techniques document is provided to assist Instructors, Coaches and Students as a training aid in the development of good soaring skills. Instructors
Gait Analyser. Description of Walking Performance
 Gait Analyser Description of Walking Performance This brochure will help you to understand clearly the parameters described in the report of the Gait Analyser, provide you with tips to implement the walking
Gait Analyser Description of Walking Performance This brochure will help you to understand clearly the parameters described in the report of the Gait Analyser, provide you with tips to implement the walking
The Mechanics of Modern BREASTSTROKE Swimming Dr Ralph Richards
 The Mechanics of Modern BREASTSTROKE Swimming Dr Ralph Richards Breaststroke is the least efficient of the four competition strokes because a large amount of water resistance is created due to body position
The Mechanics of Modern BREASTSTROKE Swimming Dr Ralph Richards Breaststroke is the least efficient of the four competition strokes because a large amount of water resistance is created due to body position
Ripple Tank Exploring the Properties of Waves Using a Ripple Tank
 Exploring the Properties of Waves Using a The ripple tank is a shallow, glass-bottomed container that is filled with water to a depth of 1 or 2 centimeters. There is a light source that is placed above
Exploring the Properties of Waves Using a The ripple tank is a shallow, glass-bottomed container that is filled with water to a depth of 1 or 2 centimeters. There is a light source that is placed above
Chapter 36. Table of Contents. Section 1 Phylum Arthropoda. Section 2 Subphylum Crustacea. Section 3 Subphylum Chelicerata and Myriapoda.
 Arthropods Table of Contents Section 1 Phylum Arthropoda Section 2 Subphylum Crustacea Section 3 Subphylum Chelicerata and Myriapoda Section 1 Phylum Arthropoda Objectives Describe the distinguishing characteristics
Arthropods Table of Contents Section 1 Phylum Arthropoda Section 2 Subphylum Crustacea Section 3 Subphylum Chelicerata and Myriapoda Section 1 Phylum Arthropoda Objectives Describe the distinguishing characteristics
Equine Cannon Angle System
 Equine Cannon System How to interpret the results December 2010 Page 1 of 14 Table of Contents Introduction... 3 The Sagittal Plane... 4 The Coronal Plane... 5 Results Format... 6 How to Interpret the
Equine Cannon System How to interpret the results December 2010 Page 1 of 14 Table of Contents Introduction... 3 The Sagittal Plane... 4 The Coronal Plane... 5 Results Format... 6 How to Interpret the
Wave Motion. interference destructive interferecne constructive interference in phase. out of phase standing wave antinodes resonant frequencies
 Wave Motion Vocabulary mechanical waves pulse continuous periodic wave amplitude period wavelength period wave velocity phase transverse wave longitudinal wave intensity displacement amplitude phase velocity
Wave Motion Vocabulary mechanical waves pulse continuous periodic wave amplitude period wavelength period wave velocity phase transverse wave longitudinal wave intensity displacement amplitude phase velocity
Technical Note. Determining the surface tension of liquids by measurements on pendant drops
 Technical Note Pendant Drop Measurements Technical note: TN316e Industry section: all Author: FT, TW Date: 12/2010 Method: Drop Shape Analyzer DSA100 Keywords: Methods, surface tension, interfacial tension,
Technical Note Pendant Drop Measurements Technical note: TN316e Industry section: all Author: FT, TW Date: 12/2010 Method: Drop Shape Analyzer DSA100 Keywords: Methods, surface tension, interfacial tension,
Timing 1. The hips and shoulders rotate at the end of the catch to assist in the acceleration of the stroke.
 Freestyle Body Position 1. Head position should be comfortable at a 45º angle with the water line slightly above the goggles. 2. Hips and shoulders should remain high in the water with a straight line
Freestyle Body Position 1. Head position should be comfortable at a 45º angle with the water line slightly above the goggles. 2. Hips and shoulders should remain high in the water with a straight line
In this course you will learn the following
 Module 11 : Example study of robots Lecture 40 : NATARAJ a case study of a 6-legged robot Objectives In this course you will learn the following Mobile Robots Legged Robots Nataraj Robot Nataraj Development
Module 11 : Example study of robots Lecture 40 : NATARAJ a case study of a 6-legged robot Objectives In this course you will learn the following Mobile Robots Legged Robots Nataraj Robot Nataraj Development
How to Make, Interpret and Use a Simple Plot
 How to Make, Interpret and Use a Simple Plot A few of the students in ASTR 101 have limited mathematics or science backgrounds, with the result that they are sometimes not sure about how to make plots
How to Make, Interpret and Use a Simple Plot A few of the students in ASTR 101 have limited mathematics or science backgrounds, with the result that they are sometimes not sure about how to make plots
Breaking Down the Approach
 Breaking Down the Approach Written by Andre Christopher Gonzalez Sunday, July 31, 2005 One of the biggest weaknesses of the two-legged approach is the inability of the athlete to transfer horizontal momentum
Breaking Down the Approach Written by Andre Christopher Gonzalez Sunday, July 31, 2005 One of the biggest weaknesses of the two-legged approach is the inability of the athlete to transfer horizontal momentum
Tropical Marine Invertebrates. Phylum Arthropoda. Phylum ARTHROPODA CAS BI 569
 Tropical Marine Invertebrates CAS BI 569 Phylum ARTHROPODA by John R. Finnerty Phylum Arthropoda Well over 1 million described extant species Four critical bodyplan features. Chitinous exoskeleton Segmented
Tropical Marine Invertebrates CAS BI 569 Phylum ARTHROPODA by John R. Finnerty Phylum Arthropoda Well over 1 million described extant species Four critical bodyplan features. Chitinous exoskeleton Segmented
Contribution to the morphology of the third-instar larvae of Laccophilus poecilus KLUG (Coleoptera: Dytiscidae)
") Genus Vol. 15(1): 31-36 Wroc³aw, 30 III 2004 Contribution to the morphology of the third-instar larvae of Laccophilus poecilus KLUG (Coleoptera: Dytiscidae) EUGENIUSZ BIESIADKA and IWONA KA KAŹMIERSKA
Genus Vol. 15(1): 31-36 Wroc³aw, 30 III 2004 Contribution to the morphology of the third-instar larvae of Laccophilus poecilus KLUG (Coleoptera: Dytiscidae) EUGENIUSZ BIESIADKA and IWONA KA KAŹMIERSKA
Ma Long's forehand Power Looping Backspin Power looping backspin is the most common attacking method, and it is one of the most powerful forehand
 Ma Long s Technique Ma Long, the latest men-single winner in Asian games, has beaten lots of top players in the world. Characteristic of his game is obvious - very good at initiating attack, fast and fierce,
Ma Long s Technique Ma Long, the latest men-single winner in Asian games, has beaten lots of top players in the world. Characteristic of his game is obvious - very good at initiating attack, fast and fierce,
Gait. Kinesiology RHS 341 Lecture 12 Dr. Einas Al-Eisa
 Gait Kinesiology RHS 341 Lecture 12 Dr. Einas Al-Eisa Definitions Locomotion = the act of moving from one place to the other Gait = the manner of walking Definitions Walking = a smooth, highly coordinated,
Gait Kinesiology RHS 341 Lecture 12 Dr. Einas Al-Eisa Definitions Locomotion = the act of moving from one place to the other Gait = the manner of walking Definitions Walking = a smooth, highly coordinated,
Slide 1. Slide 1. Next. 5:30:08 AM
 Slide 1 Slide 1 http://www3.utep.edu/leb/mosquito/larvslide1.htm10/27/2004 5:30:08 AM Slide 1 Slide 2 Recognition that the specimens are mosquito larvae is a prerequisite to identification of the genera.
Slide 1 Slide 1 http://www3.utep.edu/leb/mosquito/larvslide1.htm10/27/2004 5:30:08 AM Slide 1 Slide 2 Recognition that the specimens are mosquito larvae is a prerequisite to identification of the genera.
CHANGES OF INTERNAL HYDROSTATIC PRESSURE AND BODY SHAPE IN ACANTHOCEPHALUS RANAE
 J. Exp. Biol. (1966), 45, 197-204 197 With 5 text-figures Printed in Great Britain CHANGES OF INTERNAL HYDROSTATIC PRESSURE AND BODY SHAPE IN ACANTHOCEPHALUS RANAE BY R. A. HAMMOND Department of Zoology,
J. Exp. Biol. (1966), 45, 197-204 197 With 5 text-figures Printed in Great Britain CHANGES OF INTERNAL HYDROSTATIC PRESSURE AND BODY SHAPE IN ACANTHOCEPHALUS RANAE BY R. A. HAMMOND Department of Zoology,
THE AIRCRAFT IN FLIGHT Issue /07/12
 1 INTRODUCTION This series of tutorials for the CIX VFR Club are based on real world training. Each document focuses on a small part only of the necessary skills required to fly a light aircraft, and by
1 INTRODUCTION This series of tutorials for the CIX VFR Club are based on real world training. Each document focuses on a small part only of the necessary skills required to fly a light aircraft, and by
Designing a Model Rocket
 Designing a Model Rocket Design Components In the following pages we are going to look at the design requirements for a stable single stage model rocket. From the diagram here you can see that the rocket
Designing a Model Rocket Design Components In the following pages we are going to look at the design requirements for a stable single stage model rocket. From the diagram here you can see that the rocket
Equine Results Interpretation Guide For Cannon Angles May 2013
 Equine Results Interpretation Guide For Cannon Angles May 2013 Page 1 of 20 Table of Contents 1. Introduction... 3 2. Options for all plots... 7 3. Tables of data... 8 4. Gaits... 10 5. Walk... 10 6. Trot...
Equine Results Interpretation Guide For Cannon Angles May 2013 Page 1 of 20 Table of Contents 1. Introduction... 3 2. Options for all plots... 7 3. Tables of data... 8 4. Gaits... 10 5. Walk... 10 6. Trot...
Aerodynamic Terms. Angle of attack is the angle between the relative wind and the wing chord line. [Figure 2-2] Leading edge. Upper camber.
![Aerodynamic Terms. Angle of attack is the angle between the relative wind and the wing chord line. [Figure 2-2] Leading edge. Upper camber.](/thumbs/82/86661300.jpg "Aerodynamic Terms. Angle of attack is the angle between the relative wind and the wing chord line. [Figure 2-2] Leading edge. Upper camber.") Chapters 2 and 3 of the Pilot s Handbook of Aeronautical Knowledge (FAA-H-8083-25) apply to powered parachutes and are a prerequisite to reading this book. This chapter will focus on the aerodynamic fundamentals
Chapters 2 and 3 of the Pilot s Handbook of Aeronautical Knowledge (FAA-H-8083-25) apply to powered parachutes and are a prerequisite to reading this book. This chapter will focus on the aerodynamic fundamentals
EXSC 408L Fall '03 Problem Set #2 Linear Motion. Linear Motion
 Problems: 1. Once you have recorded the calibration frame for a data collection, why is it important to make sure the camera does not shut off? hat happens if the camera automatically shuts off after being
Problems: 1. Once you have recorded the calibration frame for a data collection, why is it important to make sure the camera does not shut off? hat happens if the camera automatically shuts off after being
An investigation of kinematic and kinetic variables for the description of prosthetic gait using the ENOCH system
 An investigation of kinematic and kinetic variables for the description of prosthetic gait using the ENOCH system K. OBERG and H. LANSHAMMAR* Amputee Training and Research Unit, University Hospital, Fack,
An investigation of kinematic and kinetic variables for the description of prosthetic gait using the ENOCH system K. OBERG and H. LANSHAMMAR* Amputee Training and Research Unit, University Hospital, Fack,
The technique of reciprocal walking using the hip guidance orthosis (hgo) with crutches
 with crutches") The technique of reciprocal walking using the hip guidance orthosis (hgo) with crutches P. B. BUTLER, R. E. MAJOR and J. H. PATRICK Orthotic Research and Locomotor Assessment Unit, The Robert Jones and
The technique of reciprocal walking using the hip guidance orthosis (hgo) with crutches P. B. BUTLER, R. E. MAJOR and J. H. PATRICK Orthotic Research and Locomotor Assessment Unit, The Robert Jones and
ZIPWAKE DYNAMIC TRIM CONTROL SYSTEM OUTLINE OF OPERATING PRINCIPLES BEHIND THE AUTOMATIC MOTION CONTROL FEATURES
 ZIPWAKE DYNAMIC TRIM CONTROL SYSTEM OUTLINE OF OPERATING PRINCIPLES BEHIND THE AUTOMATIC MOTION CONTROL FEATURES TABLE OF CONTENTS 1 INTRODUCTION 3 2 SYSTEM COMPONENTS 3 3 PITCH AND ROLL ANGLES 4 4 AUTOMATIC
ZIPWAKE DYNAMIC TRIM CONTROL SYSTEM OUTLINE OF OPERATING PRINCIPLES BEHIND THE AUTOMATIC MOTION CONTROL FEATURES TABLE OF CONTENTS 1 INTRODUCTION 3 2 SYSTEM COMPONENTS 3 3 PITCH AND ROLL ANGLES 4 4 AUTOMATIC
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission.
 Walking and Running Author(s): R. McNeill Alexander Source: The Mathematical Gazette, Vol. 80, No. 488 (Jul., 1996), pp. 262-266 Published by: The Mathematical Association Stable URL: http://www.jstor.org/stable/3619558
Walking and Running Author(s): R. McNeill Alexander Source: The Mathematical Gazette, Vol. 80, No. 488 (Jul., 1996), pp. 262-266 Published by: The Mathematical Association Stable URL: http://www.jstor.org/stable/3619558
Components of the 400m Hurdles
 Components of the 400m Hurdles By Ray Boyd NATIONAL 400M HURDLE COACH-AUSTRALIA FROM: Modern Athlete and Coach The 400m hurdles race can be broken up into four stages: Start to First Hurdle Movement Across
Components of the 400m Hurdles By Ray Boyd NATIONAL 400M HURDLE COACH-AUSTRALIA FROM: Modern Athlete and Coach The 400m hurdles race can be broken up into four stages: Start to First Hurdle Movement Across
Playing the game. The singles game
 Playing the game Before it is possible to know what is involved in playing a game it is perhaps necessary to have some idea of what is meant by the word game. A simple explanation is that a badminton game
Playing the game Before it is possible to know what is involved in playing a game it is perhaps necessary to have some idea of what is meant by the word game. A simple explanation is that a badminton game
COMPARISON STUDY BETWEEN THE EFFICIENY OF THE START TECHNIQUES IN THE ROMANIAN COMPETITIVE SWIMMING
 Bulletin of the Transilvania University of Braşov Series IX: Sciences of Human Kinetics Vol. 6 (55) No. 1 2013 COMPARISON STUDY BETWEEN THE EFFICIENY OF THE START TECHNIQUES IN THE ROMANIAN COMPETITIVE
Bulletin of the Transilvania University of Braşov Series IX: Sciences of Human Kinetics Vol. 6 (55) No. 1 2013 COMPARISON STUDY BETWEEN THE EFFICIENY OF THE START TECHNIQUES IN THE ROMANIAN COMPETITIVE
Michigan Science Olympiad Entomology Event. Coaches Meeting Dec. 7, 2013
 Michigan Science Olympiad 2014 Entomology Event Coaches Meeting Dec. 7, 2013 Entomology 2014 Test Description Test Date: April 26, 2014 Location: TBD Duration: 50 minutes Supervisor: Amanda Lorenz, Graduate
Michigan Science Olympiad 2014 Entomology Event Coaches Meeting Dec. 7, 2013 Entomology 2014 Test Description Test Date: April 26, 2014 Location: TBD Duration: 50 minutes Supervisor: Amanda Lorenz, Graduate
Advanced Hydraulics Prof. Dr. Suresh A. Kartha Department of Civil Engineering Indian Institute of Technology, Guwahati
 Advanced Hydraulics Prof. Dr. Suresh A. Kartha Department of Civil Engineering Indian Institute of Technology, Guwahati Module - 4 Hydraulics Jumps Lecture - 4 Features of Hydraulic Jumps (Refer Slide
Advanced Hydraulics Prof. Dr. Suresh A. Kartha Department of Civil Engineering Indian Institute of Technology, Guwahati Module - 4 Hydraulics Jumps Lecture - 4 Features of Hydraulic Jumps (Refer Slide
Geezer Senior Focus - What?
 Geezer Senior Focus - What? Seniors are a very discerning group of diverse individuals who are quite adept at recognizing balderdash. This means that whoever coaches seniors needs to be very knowledgeable
Geezer Senior Focus - What? Seniors are a very discerning group of diverse individuals who are quite adept at recognizing balderdash. This means that whoever coaches seniors needs to be very knowledgeable
STUDIES IN ANIMAL LOCOMOTION
 STUDIES IN ANIMAL LOCOMOTION VIII. THE KINETICS OF LOCOMOTION OF NEREIS DIVERSICOLOR BY J. GRAY Zoological Laboratory, Cambridge (Received 30 March 1938) (With One Plate and Eight Text-figures) THE locomotory
STUDIES IN ANIMAL LOCOMOTION VIII. THE KINETICS OF LOCOMOTION OF NEREIS DIVERSICOLOR BY J. GRAY Zoological Laboratory, Cambridge (Received 30 March 1938) (With One Plate and Eight Text-figures) THE locomotory
Chapters 25: Waves. f = 1 T. v =!f. Text: Chapter 25 Think and Explain: 1-10 Think and Solve: 1-4
 Text: Chapter 25 Think and Explain: 1-10 Think and Solve: 1-4 Chapters 25: Waves NAME: Vocabulary: wave, pulse, oscillation, amplitude, wavelength, wave speed, frequency, period, interference, constructive,
Text: Chapter 25 Think and Explain: 1-10 Think and Solve: 1-4 Chapters 25: Waves NAME: Vocabulary: wave, pulse, oscillation, amplitude, wavelength, wave speed, frequency, period, interference, constructive,
Respiratory Pulmonary Ventilation
 Respiratory Pulmonary Ventilation Pulmonary Ventilation Pulmonary ventilation is the act of breathing and the first step in the respiratory process. Pulmonary ventilation brings in air with a new supply
Respiratory Pulmonary Ventilation Pulmonary Ventilation Pulmonary ventilation is the act of breathing and the first step in the respiratory process. Pulmonary ventilation brings in air with a new supply
THE INITIAL STAGE THE FINAL STAGE
 THE JAVELIN RUN-UP By Hans Torim A detailed description of the author s views on the javelin run-up from the initial stages to the impulse stride and the pre-delivery position. The article is a slightly
THE JAVELIN RUN-UP By Hans Torim A detailed description of the author s views on the javelin run-up from the initial stages to the impulse stride and the pre-delivery position. The article is a slightly
THE BRIDGE COLLAPSED IN NOVEMBER 1940 AFTER 4 MONTHS OF ITS OPENING TO TRAFFIC!
 OUTLINE TACOMA NARROWS BRIDGE FLOW REGIME PAST A CYLINDER VORTEX SHEDDING MODES OF VORTEX SHEDDING PARALLEL & OBLIQUE FLOW PAST A SPHERE AND A CUBE SUMMARY TACOMA NARROWS BRIDGE, USA THE BRIDGE COLLAPSED
OUTLINE TACOMA NARROWS BRIDGE FLOW REGIME PAST A CYLINDER VORTEX SHEDDING MODES OF VORTEX SHEDDING PARALLEL & OBLIQUE FLOW PAST A SPHERE AND A CUBE SUMMARY TACOMA NARROWS BRIDGE, USA THE BRIDGE COLLAPSED
Coaching the Hurdles
 Coaching the Hurdles Monica Gary, Sprints & Hurdles Coach Purdue University Important components to consider in hurdle training: a. Rhythm for the hurdler is the primary concern for the coach -short rhythm
Coaching the Hurdles Monica Gary, Sprints & Hurdles Coach Purdue University Important components to consider in hurdle training: a. Rhythm for the hurdler is the primary concern for the coach -short rhythm
Rules of Hurdling. Distance Between Hurdles
 The Hurdle Events Introduction Brief discussion of rules, safety practices, and talent demands for the hurdles. Examine technical and training considerations for the hurdle events. 100 Meter Hurdles for
The Hurdle Events Introduction Brief discussion of rules, safety practices, and talent demands for the hurdles. Examine technical and training considerations for the hurdle events. 100 Meter Hurdles for
THE IMPULSE-STEP IN THE JAVELIN THROW
 THE IMPULSE-STEP IN THE JAVELIN THROW Terseus Liebenberg North-West University Potchefstroom South Africa The most important biomechanical principle ultimately determining throwing distance is the release
THE IMPULSE-STEP IN THE JAVELIN THROW Terseus Liebenberg North-West University Potchefstroom South Africa The most important biomechanical principle ultimately determining throwing distance is the release
Harmonic Motion: Pendulums Student Version
 Harmonic Motion: Pendulums Student Version In this lab you will set up a pendulum using rulers, string, and small weights and measure how different variables affect the period of the pendulum. You will
Harmonic Motion: Pendulums Student Version In this lab you will set up a pendulum using rulers, string, and small weights and measure how different variables affect the period of the pendulum. You will
YIN&YANG ENERGY LINES
 YIN&YANG ENERGY LINES Karate technique is a combination of intentional creation and distribution of the energy - a physical movement that enables human body the use of its full potential at the time. In
YIN&YANG ENERGY LINES Karate technique is a combination of intentional creation and distribution of the energy - a physical movement that enables human body the use of its full potential at the time. In
2008 Study Update, Part 3 By Dr. Ed Ashby
 2008 Study Update, Part 3 By Dr. Ed Ashby In Parts 1 and 2 we looked at the results of the 40# bow's Heavy Bone Threshold testing and compared those results to that shown by 'commonly used' arrows from
2008 Study Update, Part 3 By Dr. Ed Ashby In Parts 1 and 2 we looked at the results of the 40# bow's Heavy Bone Threshold testing and compared those results to that shown by 'commonly used' arrows from
SIMULTANEOUS RECORDINGS OF VELOCITY AND VIDEO DURING SWIMMING
 Portuguese Journal of Sport Sciences. 6:supl. 2, 32-35, 2006 SIMULTANEOUS RECORDINGS OF VELOCITY AND VIDEO DURING SWIMMING Albert B. Craig 1, Budd Termin2, and David R. Pendergast 2 1University of Rochester,
Portuguese Journal of Sport Sciences. 6:supl. 2, 32-35, 2006 SIMULTANEOUS RECORDINGS OF VELOCITY AND VIDEO DURING SWIMMING Albert B. Craig 1, Budd Termin2, and David R. Pendergast 2 1University of Rochester,
(fig. 3) must be at the same temperature as the water in this chamber CALORIMETRIC STUDIES OF THE EXTREMITIES
 must be at the same temperature as the water in this chamber CALORIMETRIC STUDIES OF THE EXTREMITIES") CALORIMETRIC STUDIES OF THE EXTREMITIES II. EXPERIMENTAL APPARATUS AND PROCEDURES' By ROY KEGERREIS (Received for publication July 1, 1926) The calorimeter used in these experiments is a modification of
CALORIMETRIC STUDIES OF THE EXTREMITIES II. EXPERIMENTAL APPARATUS AND PROCEDURES' By ROY KEGERREIS (Received for publication July 1, 1926) The calorimeter used in these experiments is a modification of
Decentralized Autonomous Control of a Myriapod Locomotion Robot
 Decentralized utonomous Control of a Myriapod Locomotion Robot hmet Onat Sabanci University, Turkey onat@sabanciuniv.edu Kazuo Tsuchiya Kyoto University, Japan tsuchiya@kuaero.kyoto-u.ac.jp Katsuyoshi
Decentralized utonomous Control of a Myriapod Locomotion Robot hmet Onat Sabanci University, Turkey onat@sabanciuniv.edu Kazuo Tsuchiya Kyoto University, Japan tsuchiya@kuaero.kyoto-u.ac.jp Katsuyoshi
The Starting Point. Prosthetic Alignment in the Transtibial Amputee. Outline. COM Motion in the Coronal Plane
 Prosthetic Alignment in the Transtibial Amputee The Starting Point David C. Morgenroth, MD, Department of Rehabilitation Medicine University of Washington VAPSHCS Outline COM Motion in the Coronal Plane
Prosthetic Alignment in the Transtibial Amputee The Starting Point David C. Morgenroth, MD, Department of Rehabilitation Medicine University of Washington VAPSHCS Outline COM Motion in the Coronal Plane
Chapter 11 Waves. Waves transport energy without transporting matter. The intensity is the average power per unit area. It is measured in W/m 2.
 Energy can be transported by particles or waves: Chapter 11 Waves A wave is characterized as some sort of disturbance that travels away from a source. The key difference between particles and waves is
Energy can be transported by particles or waves: Chapter 11 Waves A wave is characterized as some sort of disturbance that travels away from a source. The key difference between particles and waves is
2008 Excellence in Mathematics Contest Team Project B. School Name: Group Members:
 2008 Excellence in Mathematics Contest Team Project B School Name: Group Members: Lassie - TEAM PROJECT B 2008 Excellence in Mathematics Contest The Team Project is a group activity in which the students
2008 Excellence in Mathematics Contest Team Project B School Name: Group Members: Lassie - TEAM PROJECT B 2008 Excellence in Mathematics Contest The Team Project is a group activity in which the students
WESTERN DRESSAGE GLOSSARY
 WESTERN DRESSAGE GLOSSARY GAITS Any of the rhythms or footfalls of the horse. Gaits demonstrated in Western Dressage include: walk, jog, and lope. Freedom and regularity should be maintained in all three
WESTERN DRESSAGE GLOSSARY GAITS Any of the rhythms or footfalls of the horse. Gaits demonstrated in Western Dressage include: walk, jog, and lope. Freedom and regularity should be maintained in all three
PARA-BADMINTON WHEELCHAIR DEVELOPMENT
 PARA-BADMINTON WHEELCHAIR DEVELOPMENT Para-Badminton Wheelchair Development Introduction The use of the arms for both movement and hitting technique are the main challenges in the wheelchair sports class.
PARA-BADMINTON WHEELCHAIR DEVELOPMENT Para-Badminton Wheelchair Development Introduction The use of the arms for both movement and hitting technique are the main challenges in the wheelchair sports class.
Waves. harmonic wave wave equation one dimensional wave equation principle of wave fronts plane waves law of reflection
 Waves Vocabulary mechanical wave pulse continuous periodic wave amplitude wavelength period frequency wave velocity phase transverse wave longitudinal wave intensity displacement wave number phase velocity
Waves Vocabulary mechanical wave pulse continuous periodic wave amplitude wavelength period frequency wave velocity phase transverse wave longitudinal wave intensity displacement wave number phase velocity
Calculation of Trail Usage from Counter Data
 1. Introduction 1 Calculation of Trail Usage from Counter Data 1/17/17 Stephen Martin, Ph.D. Automatic counters are used on trails to measure how many people are using the trail. A fundamental question
1. Introduction 1 Calculation of Trail Usage from Counter Data 1/17/17 Stephen Martin, Ph.D. Automatic counters are used on trails to measure how many people are using the trail. A fundamental question
USA Track & Field Heptathlon Summit- November
 USA Track & Field Heptathlon Summit- November 1994 1 I. Technical considerations in the sprint hurdles Practical Biomechanics For the 100m Hurdles By Gary Winckler University of Illinois A. General flow
USA Track & Field Heptathlon Summit- November 1994 1 I. Technical considerations in the sprint hurdles Practical Biomechanics For the 100m Hurdles By Gary Winckler University of Illinois A. General flow
Walking Tall: Mobility Drills for Seniors
 Walking Tall: Mobility Drills for Seniors What is Functional Mobility Training? Selecting exercises that improve the foundation for movement Working in multiple planes Teaching reaction time, decision
Walking Tall: Mobility Drills for Seniors What is Functional Mobility Training? Selecting exercises that improve the foundation for movement Working in multiple planes Teaching reaction time, decision
Ascent to Altitude After Diving
 Ascent to Altitude After Diving On many occasions, divers have a need to ascend to a higher altitude after diving, and they need guidance on how long they need to wait before doing so. The reason they
Ascent to Altitude After Diving On many occasions, divers have a need to ascend to a higher altitude after diving, and they need guidance on how long they need to wait before doing so. The reason they
Figure 1. The distance the train travels between A and B is not the same as the displacement of the train.
 THE DISTANCE-TIME RELATIONSHIP Q1. A train travels from town A to town B. Figure 1 shows the route taken by the train. Figure 1 has been drawn to scale. Figure 1 (a) The distance the train travels between
THE DISTANCE-TIME RELATIONSHIP Q1. A train travels from town A to town B. Figure 1 shows the route taken by the train. Figure 1 has been drawn to scale. Figure 1 (a) The distance the train travels between
Chapter # 08 Waves. [WAVES] Chapter # 08
![Chapter # 08 Waves. [WAVES] Chapter # 08](/thumbs/94/121079132.jpg "Chapter # 08 Waves. [WAVES] Chapter # 08") Chapter # 08 Waves Q2) Write short answers of the following questions. i) What is the difference between progressive and stationary waves? Answer: Progressive Waves 1 Progressive waves are the result of
Chapter # 08 Waves Q2) Write short answers of the following questions. i) What is the difference between progressive and stationary waves? Answer: Progressive Waves 1 Progressive waves are the result of
TECHNIQUE AND METHODOLOGY OF TRAINING IN SWIMMING CRAWL
 Selim Alili Tetovo, Macedonia UDK: 797.212.4 TECHNIQUE AND METHODOLOGY OF TRAINING IN SWIMMING CRAWL 1. INTRODUCTION As a father of modern crawl is considered Australia. The main feature of this mode of
Selim Alili Tetovo, Macedonia UDK: 797.212.4 TECHNIQUE AND METHODOLOGY OF TRAINING IN SWIMMING CRAWL 1. INTRODUCTION As a father of modern crawl is considered Australia. The main feature of this mode of
plethysmographic methods that when the subject was pinched on the upper
 24 J. Physiol. (I95I) II2, 24-2I 6I2.I5.6II.976 THE DECREASE IN HAND BLOOD FLOW FOLLOWING INFLATION OF AN ARTERIAL OCCLUSION CUFF ON THE OPPOSITE ARM BY IAN C. RODDIE From the Department of Physiology,
24 J. Physiol. (I95I) II2, 24-2I 6I2.I5.6II.976 THE DECREASE IN HAND BLOOD FLOW FOLLOWING INFLATION OF AN ARTERIAL OCCLUSION CUFF ON THE OPPOSITE ARM BY IAN C. RODDIE From the Department of Physiology,
GEA FOR ADVANCED STRUCTURAL DYNAMIC ANALYSIS
 SMART SOLUTIONS FOR VIBRATION MONITORING GEA FOR ADVANCED STRUCTURAL DYNAMIC ANALYSIS ANALYSIS OF CIVIL STRUCTURES - EXPO MERLATA PEDESTRIAN BRIDGE ABSTRACT Civil structures and in particular bridges and
SMART SOLUTIONS FOR VIBRATION MONITORING GEA FOR ADVANCED STRUCTURAL DYNAMIC ANALYSIS ANALYSIS OF CIVIL STRUCTURES - EXPO MERLATA PEDESTRIAN BRIDGE ABSTRACT Civil structures and in particular bridges and
Wave phenomena in a ripple tank
 Wave phenomena in a ripple tank LEP Related topics Generation of surface waves, propagation of surface waves, reflection of waves, refraction of waves, Doppler Effect. Principle Water waves are generated
Wave phenomena in a ripple tank LEP Related topics Generation of surface waves, propagation of surface waves, reflection of waves, refraction of waves, Doppler Effect. Principle Water waves are generated
Normal and Abnormal Gait
 Normal and Abnormal Gait Adrielle Fry, MD EvergreenHealth, Division of Sport and Spine University of Washington Board Review Course March 6, 2017 What are we going to cover? Definitions and key concepts
Normal and Abnormal Gait Adrielle Fry, MD EvergreenHealth, Division of Sport and Spine University of Washington Board Review Course March 6, 2017 What are we going to cover? Definitions and key concepts
(From the Physiological and Biochemical Laboratories, Cambridge.)
") CREATINE FORMATION DURING TONIC MUSCLE CONTRACTION. BY K. UYENO AND T. MITSUDA. (From the Physiological and Biochemical Laboratories, Cambridge.) Creatine of the amphibition muscles in the breeding season.
CREATINE FORMATION DURING TONIC MUSCLE CONTRACTION. BY K. UYENO AND T. MITSUDA. (From the Physiological and Biochemical Laboratories, Cambridge.) Creatine of the amphibition muscles in the breeding season.
Performance tests and explanation of evaluation criteria IBOP RIDING TEST
 Performance tests and explanation of evaluation criteria IBOP RIDING TEST 1. A X - C Enter at working trot C Follow left track 2. A Large circle 3. H X F Change hands, and at X transition to working walk
Performance tests and explanation of evaluation criteria IBOP RIDING TEST 1. A X - C Enter at working trot C Follow left track 2. A Large circle 3. H X F Change hands, and at X transition to working walk
Lesson: Pitch Trim. Materials / Equipment Publications o Flight Training Manual for Gliders (Holtz) Lesson 4.4 Using the Trim Control.
 Lesson 4.4 Using the Trim Control.") 11/18/2015 Pitch Trim Page 1 Lesson: Pitch Trim Objectives: o Knowledge o An understanding of the aerodynamics related to longitudinal (pitch) stability o Skill o Use of the pitch trim system to control
11/18/2015 Pitch Trim Page 1 Lesson: Pitch Trim Objectives: o Knowledge o An understanding of the aerodynamics related to longitudinal (pitch) stability o Skill o Use of the pitch trim system to control
1/10/2013. Musculoskeletal Integration. What forces is a body subjected to on land? What forces is a body subjected to on land?
 Topic : Support & Locomotion on Land What forces is the body subjected to on land? Balance Posture Gait How is the vertebral column involved in locomotion? Support Movement How are the limbs involved in
Topic : Support & Locomotion on Land What forces is the body subjected to on land? Balance Posture Gait How is the vertebral column involved in locomotion? Support Movement How are the limbs involved in
Invertebrate Notes. Arthropoda. Subphylum: Cheliceratahorseshoe. crabs, spiders, scorpions, mites, & ticks
 Invertebrate Notes Invertebrate Phyla: Trilobita-- trilobites (extinct) Arthropoda Cheliceratahorseshoe crabs, spiders, scorpions, mites, & ticks Mandibulata-- crustaceans, insects, millipedes, centipedes
Invertebrate Notes Invertebrate Phyla: Trilobita-- trilobites (extinct) Arthropoda Cheliceratahorseshoe crabs, spiders, scorpions, mites, & ticks Mandibulata-- crustaceans, insects, millipedes, centipedes
Swimming Stroke Mechanics
 Swimming Stroke Mechanics What we continue to learn from High-speed Videography and Biomechanical Motion Analysis Jan Prins, Ph.D. Aquatic Research Laboratory University of Hawaii Swimming Biomechanics,
Swimming Stroke Mechanics What we continue to learn from High-speed Videography and Biomechanical Motion Analysis Jan Prins, Ph.D. Aquatic Research Laboratory University of Hawaii Swimming Biomechanics,
Pendant Drop Measurements
 KRÜSS pplication Note TN316d Page 1 Pendant Drop Measurements pplication note: TN316d Industry section: all uthor: Dr. Tobias Winkler Date: December 2010 Method: Drop Shape nalysis System DS100 Drop Shape
KRÜSS pplication Note TN316d Page 1 Pendant Drop Measurements pplication note: TN316d Industry section: all uthor: Dr. Tobias Winkler Date: December 2010 Method: Drop Shape nalysis System DS100 Drop Shape
Harmonic Motion: Pendulums
 Harmonic Motion: Pendulums In this lab you will set up a pendulum using rulers, string, and small weights and measure how different variables affect the period of the pendulum. You will also use the concept
Harmonic Motion: Pendulums In this lab you will set up a pendulum using rulers, string, and small weights and measure how different variables affect the period of the pendulum. You will also use the concept
Traveling Waves vs. Standing Waves
 The Physics Classroom» Physics Tutorial» Waves» Traveling Waves vs. Standing Waves Waves - Lesson 4 - Standing Waves Traveling Waves vs. Standing Waves Traveling Waves vs. Standing Waves Formation of Standing
The Physics Classroom» Physics Tutorial» Waves» Traveling Waves vs. Standing Waves Waves - Lesson 4 - Standing Waves Traveling Waves vs. Standing Waves Traveling Waves vs. Standing Waves Formation of Standing
THE DEVELOPMENT OF SPEED:
 THE DEVELOPMENT OF SPEED: BEFORE YOU HOP ON THAT TREADMILL.. By Jason Shea, M.S., C.S.C.S, PES Doing a quick internet search for treadmill manufacturers, one can see what a lucrative business it is to
THE DEVELOPMENT OF SPEED: BEFORE YOU HOP ON THAT TREADMILL.. By Jason Shea, M.S., C.S.C.S, PES Doing a quick internet search for treadmill manufacturers, one can see what a lucrative business it is to
Rugby Strength Coach. Speed development guide
 Rugby Strength Coach Speed development guide Outline Why do Newton's laws of motion matter? What is speed? The technique and physical demands of speed Speed training parameters Rugby specific speed training
Rugby Strength Coach Speed development guide Outline Why do Newton's laws of motion matter? What is speed? The technique and physical demands of speed Speed training parameters Rugby specific speed training
3. GRADUALLY-VARIED FLOW (GVF) AUTUMN 2018
 AUTUMN 2018") 3. GRADUALLY-VARIED FLOW (GVF) AUTUMN 2018 3.1 Normal Flow vs Gradually-Varied Flow V 2 /2g EGL (energy grade line) Friction slope S f h Geometric slope S 0 In flow the downslope component of weight balances
3. GRADUALLY-VARIED FLOW (GVF) AUTUMN 2018 3.1 Normal Flow vs Gradually-Varied Flow V 2 /2g EGL (energy grade line) Friction slope S f h Geometric slope S 0 In flow the downslope component of weight balances
PHASE 1 WIND STUDIES REPORT
 PHASE 1 WIND STUDIES REPORT ENVIRONMENTAL STUDIES AND PRELIMINARY DESIGN FOR A SUICIDE DETERRENT SYSTEM Contract 2006-B-17 24 MAY 2007 Golden Gate Bridge Highway and Transportation District Introduction
PHASE 1 WIND STUDIES REPORT ENVIRONMENTAL STUDIES AND PRELIMINARY DESIGN FOR A SUICIDE DETERRENT SYSTEM Contract 2006-B-17 24 MAY 2007 Golden Gate Bridge Highway and Transportation District Introduction
400-Meter Hurdle Theory By Ralph Lindeman
 400-Meter Hurdle Theory By Ralph Lindeman INTRODUCTION The 400m hurdle race is arguably the most demanding of all events in the sprint-hurdle group. It requires a combination of speed-endurance and hurdling
400-Meter Hurdle Theory By Ralph Lindeman INTRODUCTION The 400m hurdle race is arguably the most demanding of all events in the sprint-hurdle group. It requires a combination of speed-endurance and hurdling
The effect of pack formation at the 2005 world orienteering championships
 The effect of pack formation at the 2005 world orienteering championships Prof. Graeme J. Ackland School of Physics, University of Edinburgh Edinburgh EH9 3JZ Abstract At the 2005 world championships there
The effect of pack formation at the 2005 world orienteering championships Prof. Graeme J. Ackland School of Physics, University of Edinburgh Edinburgh EH9 3JZ Abstract At the 2005 world championships there
Gerald D. Anderson. Education Technical Specialist
 Gerald D. Anderson Education Technical Specialist The factors which influence selection of equipment for a liquid level control loop interact significantly. Analyses of these factors and their interactions
Gerald D. Anderson Education Technical Specialist The factors which influence selection of equipment for a liquid level control loop interact significantly. Analyses of these factors and their interactions
Road Markings. Lecture Notes in Transportation Systems Engineering. Prof. Tom V. Mathew
 Road Markings Lecture Notes in Transportation Systems Engineering Prof. Tom V. Mathew 1 Overview The essential purpose of road markings is to guide and control traffic on a highway. They supplement the
Road Markings Lecture Notes in Transportation Systems Engineering Prof. Tom V. Mathew 1 Overview The essential purpose of road markings is to guide and control traffic on a highway. They supplement the
Skier Evaluation Form
 Skier Evaluation Form Instructor Alignment Stance Discipline Reference Alignments Terrain Green Blue Black Off-Piste : Side Slip w/ Stop Wedge Change-Ups Hockey Stop Hops in a Straight Run Side Slips with
Skier Evaluation Form Instructor Alignment Stance Discipline Reference Alignments Terrain Green Blue Black Off-Piste : Side Slip w/ Stop Wedge Change-Ups Hockey Stop Hops in a Straight Run Side Slips with
by Michael Young Human Performance Consulting
 by Michael Young Human Performance Consulting The high performance division of USATF commissioned research to determine what variables were most critical to success in the shot put The objective of the
by Michael Young Human Performance Consulting The high performance division of USATF commissioned research to determine what variables were most critical to success in the shot put The objective of the
ABOUT THE TEACHING OF THROWING EVENTS
 ABOUT THE TEACHING OF THROWING EVENTS By Friedheld Endemann The following address provides a general outline of principles applied to learning practices in throwing events with recommended sequences for
ABOUT THE TEACHING OF THROWING EVENTS By Friedheld Endemann The following address provides a general outline of principles applied to learning practices in throwing events with recommended sequences for
Notes Session #2. The second gravity organization system is the relationship of the feet with the ground.
 Notes Session #2 The first gravity organization system is the relationship of the head with space. The inner ear and the suboccipital muscles are critical here. The second gravity organization system is
Notes Session #2 The first gravity organization system is the relationship of the head with space. The inner ear and the suboccipital muscles are critical here. The second gravity organization system is
CAUDAL FIN AND BODY MOVEMENT IN THE PROPULSION OF SOME FISH
 J. Exp. Biol. (1963), 40. 23-56 23 With 1 plate and 21 text-figures Printed in Great Britain CAUDAL FIN AND BODY MOVEMENT IN THE PROPULSION OF SOME FISH BY RICHARD BAINBRIDGE Zoological Laboratory, Cambridge
J. Exp. Biol. (1963), 40. 23-56 23 With 1 plate and 21 text-figures Printed in Great Britain CAUDAL FIN AND BODY MOVEMENT IN THE PROPULSION OF SOME FISH BY RICHARD BAINBRIDGE Zoological Laboratory, Cambridge