Title. Issue Date Doc URL. Type. File Information. article (author version) cjfas final.
|
|
|
- Philip Wilkins
- 5 years ago
- Views:
Transcription
1 Title Comparison of the swimming ability and upstream-migr Author(s)Miyoshi, Koji; Hayashida, Kazufumi; Sakashita, Taku; Canadian journal of fisheries and aquatic sciences = Citation Issue Date Doc URL Type article (author version) File Information cjfas final.pdf Instructions for use Hokkaido University Collection of Scholarly and Aca
2 1 2 Comparison of the swimming ability and upstream-migration behavior between chum salmon and masu salmon 3 4 Koji Miyoshi, Kazufumi Hayashida, Taku Sakashita, Makoto Fujii, Hisaya Nii, 5 Katsuya Nakao, and Hiroshi Ueda 6 7 Koji Miyoshi, Kazufumi Hayashida, and Hiroshi Ueda. Division of Biosphere Science, 8 Graduate School of Environmental Science, Hokkaido University, North 9 West 9, Kita-ku, 9 Sapporo, Hokkaido , Japan. 10 Kazufumi Hayashida. Watershed Environmental Engineering Research Team, Civil 11 Engineering Research Institute for Cold Region, Hiragishi 1-3, Toyohira-ku, Sapporo, 12 Hokkaido , Japan. 13 Taku Sakashita, Makoto Fujii, Hisaya Nii, and Katsuya Nakao. Hokkaido Aquaculture 14 Promotion Corporation, Yonesato , Shiroishi-ku, Sapporo, Hokkaido , Japan. 15 Hiroshi Ueda. Field Science Center for Northern Biosphere, Hokkaido University, North 9 16 West 9, Kita-ku, Sapporo, Hokkaido , Japan address of each author: K. Miyoshi: miyoshi0503@gmail.com, K. Hayashida: 19 hayashida0109@gmail.com, T. Sakashita: sakashita@saibai.or.jp, M. Fujii: fujii@saibai.or.jp, 1
3 20 H. Nii: K. Nakao: and H. Ueda: Koji Miyoshi s present address. Abashiri Fisheries Research Institute, Fisheries Research 24 Department, Hokkaido Research Organization, Masu Ura 1-1, Abashiri, Hokkaido , 25 Japan Corresponding author: Dr. Hiroshi Ueda, Field Science Center for Northern Biosphere, 28 Hokkaido University, North 9 West 9, Kita-ku, Sapporo, Hokkaido , Japan. 29 Tel and Fax number: , address: hueda@fsc.hokudai.ac.jp 30 2
4 31 Abstract: The spawning ground of chum salmon (Oncorhynchus keta) is usually 32 located farther downriver than that of masu salmon (O. masou) in Hokkaido, Japan. To 33 compare the swimming abilities of these two species, the relationship between 34 swimming speed and oxygen consumption was compared using a swim tunnel in the 35 laboratory. Then, the upstream-migration behaviors of chum and masu salmon were 36 compared using electromyogram (EMG) telemetry at fish passages in the Toyohira 37 River, Hokkaido. In the laboratory study, the standard metabolic rate of masu salmon 38 was lower and the critical swimming speed (Ucrit) was faster than those of chum salmon. 39 In the field study, the holding time needed to recover the swimming performance 40 exceeding Ucrit at the fish passages, and the trial number needed to pass the fish 41 passages were significantly lower for masu salmon than chum salmon. These results 42 revealed that masu salmon are more adaptable to extended swimming in 43 high-water-velocity conditions than chum salmon and that masu salmon are better 44 equipped for a long distance upstream to their spawning ground than chum salmon Keywords: swimming ability; upstream-migration behavior; standard metabolic rate; 47 critical swimming speed; fish passage; chum salmon; masu salmon. 3
5 48 Introduction 49 Semelparous Pacific salmon must migrate upstream with limited energy stores to 50 reach their spawning ground, and the metabolic energy use involved in swimming 51 ability is important for their upstream migration. The relationships between metabolic 52 traits and swimming performance in salmon have been investigated previously. Gamperl 53 et al. (2002) showed that wild rainbow trout (Oncorhynchus mykiss) exhibit aerobic 54 fitness of at least one-third greater than that of hatchery-raised individuals. Lee et al. 55 (2003b) suggested that the heightened aerobic fitness might be related to natural 56 experiences during exercise. Wagner et al. (2006) investigated the differences in routine 57 and active metabolic rates for sexually maturing migratory adult sockeye salmon (O. 58 nerka) that were intercepted in the ocean and then held in either seawater or freshwater, 59 suggesting that some of the metabolic costs might change during the migration from 60 seawater to freshwater. 61 The upstream migration of salmon is energetically demanding because the fish 62 must pass a variety of natural and anthropogenic barriers, such as waterfalls, rapidly 63 flowing water, weirs, hydroelectric facilities, and ground sills in their natal rivers (Pon 64 et al. 2006; 2009a, b). Specifically anthropogenic barriers have brought serious 65 problems during the upstream migration of anadromous salmon. The upstream 4
6 66 spawning migration of sockeye salmon in the Fraser River, British Columbia, Canada, 67 has been extensively examined (Hinch et al. 1996; Hinch and Rand 1998; Hinch and 68 Bratty 2000), and many physiological data have been reported: the average swimming 69 activity of sockeye salmon passing through Hell s Gate were higher than at any other 70 location in the Fraser Canyon and non-aerobic swimming performance exceeding 71 critical swimming speed (Ucrit) had to be sustained over a long period of time. Roscoe 72 and Hinch (2010) reported that these barriers impede or hinder the spawning-migration 73 routes of salmon and result in reduced salmon populations. In response, a large number 74 of fish passages have been installed all over the world. 75 The recent rapid development of radio telemetry technology and methods make it 76 possible to observe the upstream-migration behavior of salmon (Cooke et al. 2004). 77 Fish telemetry with integrated muscle electromyogram (EMG) has been used to 78 examine the upstream migration of sockeye salmon (Hinch et al. 1996; Hinch & Bratty, ; Pon et al. 2009a, b). EMG can be used directly as an indicator of the intensity of 80 fish muscle activity and to evaluate the efficiency of fish passage in relation to the 81 swimming performance of Pacific salmon (Roscoe and Hinch 2010; Makiguchi et al ). 83 In Hokkaido, Japan, the spawning seasons of chum salmon (O. keta) and masu 5
7 84 salmon (O. masou) are nearly identical, but there are many differences in their upstream 85 spawning migrations. Masu salmon enter their natal river between April and June and 86 then migrate upstream to the spawning ground near the riverhead in September, possibly 87 triggered by the swollen autumnal river (Kato 1991; Mayama 1992). In contrast, chum 88 salmon begin upstream migration between September and October and quickly reach 89 the spawning ground in the middle reaches of the river (Salo 1991). We hypothesized 90 that differences in swimming ability might shape the different upstream-migration 91 behaviors of chum and masu salmon; however, to date, no study has examined the 92 relationship between swimming ability in the laboratory and upstream-migration 93 behavior in the field using chum and masu salmon. 94 The purposes of this study were (1) to examine the relationship between swimming 95 speed and oxygen consumption, and compare swimming abilities of chum and masu 96 salmon in a swim tunnel in the laboratory using Ucrit, standard metabolic rate (SMR), 97 active metabolic rate (AMR), and aerobic metabolic scope (AMS); (2) to investigate the 98 upstream-migration behaviors of chum and masu salmon in the field using EMG 99 telemetry to compare their behaviors at the fish passages in protection beds and 100 groundsills in the Toyohira River; and (3) to discuss their upstream spawning-migration 101 behavior in terms of metabolic energy use and swimming ability. 6
8 Materials and Methods 104 1) Swimming abilities in the laboratory 105 Experimental animals 106 Adult chum salmon (3 males and 2 females; mean ± SE: fork length: 62.1 ± 4.9 cm; 107 body weight: 1.9 ± 0.7 kg) and adult masu salmon (6 males and 5 females; fork length: ± 13.4 cm; body weight: 1.7 ± 1.4 kg) were captured using a waterwheel located 109 approximately 70 km from the Chitose River mouth in the upper reaches of the Ishikari 110 River in western Hokkaido, Japan, during their upstream spawning migration (Fig 1A). 111 Experiments were conducted at the Chitose Salmon Aquarium in September and 112 December of 2010 and Fish were individually transferred to a compact water tank 113 (L W H = m) in an artificially flowing stream using water from the 114 Chitose River. 115 Swim tunnel and swimming test protocol 116 A swim tunnel (West Japan Fluid Engineering Laboratory Co. Ltd, Nagasaki, 117 Japan) was used to measure critical swimming speed (Ucrit) and dissolved oxygen 118 consumption. Water flow was generated using a voltage-controlled motor and propeller, 119 with the voltage calibrated against the flow velocity. The swim tunnel was sealed with 7
9 120 an acrylic board so that no gas exchange occurred, and water from the Chitose River 121 was pumped into it before the Ucrit trial. The water temperature ranged from 12.0 to ºC and did not vary more than 1ºC during any experiments. 123 Fish were individually placed into the swimming chamber of the swim tunnel. In 124 all cases, fish were acclimated for 1 hour at 0.25 body lengths (BL) s -1 to minimize 125 handling effects before all Ucrit trials. After each 15-min period, the water velocity was 126 increased by an additional 0.25 BL s -1 and was maintained at the new speed for 15 min 127 until the fish became fatigued or was unable to swim against the current. After the Ucrit 128 trial was completed, body mass and fork length were measured. Flow-velocity values 129 were corrected to account for solid blocking effects (Gehrke et al. 1990), as described 130 by Bell and Terhune (1970). 131 The Ucrit (BL s -1 ) was calculated with Brett's formula (1964): 132 Ucrit = Ui + (t ti 1 ) U (1) 133 where Ui (BL s -1 ) is the highest velocity maintained for the entire swimming period; ti is 134 the prescribed swimming period (i.e., 15 min); t (min) is the amount of time spent at the 135 fatigue velocity; and U is the velocity increment (i.e., 0.25 BL s -1 ). 136 To measure oxygen consumption (MO2) during the acclimatization period and 137 during the Ucrit trials, the oxygen concentration in the swim tunnel was measured 8
10 138 throughout the trial at 1-min intervals using a multi-water-quality sensor probe (U-50; 139 Horiba Ltd., Kyoto, Japan), which was housed inside the swim tunnel. Before the fish 140 were introduced, air bubbles were removed from the swim tunnel. Prior to each Ucrit 141 trial, oxygen levels in the tunnel were reset by pumping fresh river water into the tunnel. 142 The MO2 per 15-min period for each fish was calculated by subtracting the quantity of 143 oxygen at the end from the starting value. The MO2 rate (mg O2 kg-1 h-1) for individual 144 fish during a velocity increment was calculated as: 145 MO2 = [O2] v/m (2) 146 where the rate of oxygen concentration [O2] is measured in mg O2 per l 1 h 1 ; v is the 147 water volume of the swim chamber (l); and m is the body mass of the fish (kg). MO2 148 was then standardized to a value corrected for a fish of 1 kg (MO2cor), using the 149 following allometric relationship derived by Schurmann and Steffensen (1997): 150 MO2cor =MO2meas (w w 1 cor) (1-A) (3) 151 where MO2meas is the MO2 measured during the trial; w is the mass of the fish; and wcor 152 is the standard body mass of 1 kg. A is the condition factor [weight (g) / 153 length 3 (cm) 100], calculated using the mean fork length and bodyweight for each 154 species. 155 Standard metabolic rate (SMR) is defined as the MO2 at zero activity (Fry 1971; 9
11 156 Claireaux et al. 2006; Luna-Acosta et al. 2011). Because MO2 was expected to increase 157 exponentially with swimming speed (Brett 1964), a curve was fitted for each individual, 158 according to the following equation: 159 MO2cor = SMR exp bu (4) 160 where MO2cor is the oxygen consumption obtained from (3); SMR is the standard 161 metabolic rate that corresponds to the intercept (i.e., the MO2 when U = 0 BL s 1 ); U is 162 the swimming speed (in BL s 1 ); and b is a constant. 163 The active metabolic rate (AMR) of the fish was estimated as the maximum MO2 164 recorded during the swimming test (i.e., when swimming speed was close to Ucrit), and 165 AMS (aerobic metabolic scope) was calculated as follows (Fry 1971): 166 AMS = AMR SMR (5) 167 The net cost of transport (COTnet; mgo2 kg 1 m 1 ) was calculated as follows for 168 each swimming speed: 169 COTnet =MO2net /U (6) 170 where U is the swimming speed (m h 1 ) and MO2-net is equal to the MO2 at the 171 considered swimming speed minus SMR. AMR, SMR, AMS and COTnet values were 172 determined for each fish
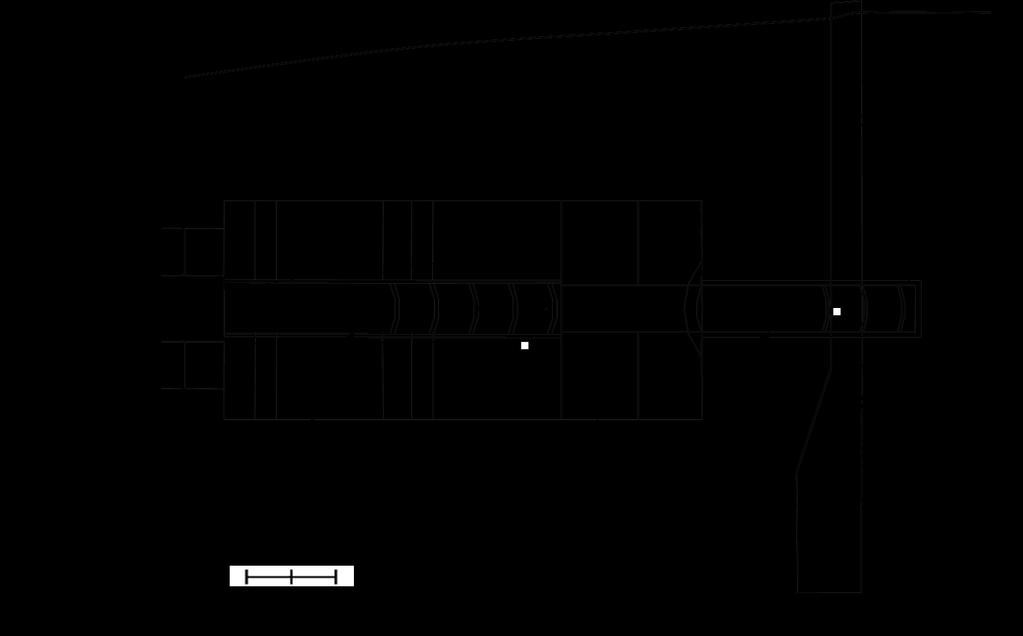
12 174 2) Upstream migration behavior in the field 175 Study area 176 The Toyohira River in Hokkaido, Japan, which flows through the city of Sapporo, 177 is a rare rapid-flow urban river and an important spawning ground for chum and masu 178 salmon. Six cross-river constructions (#1, #3-8), called groundsills, were constructed to 179 prevent the lowering of the riverbed, and fish passages were built at each groundsill (Fig B). A protection bed was also constructed at the groundsill #5, and a fish passage was 181 installed in March 2010 (Fig. 2). Makiguchi et al. (2011) reported the effects of 182 groundsills on the upstream migration of chum salmon in 2008 and 2009 using EMG 183 telemetry; however, the Toyohira River environment changed significantly after their 184 investigation; the fish passage was established in protection bed #5 in 2010, and a 185 severe typhoon hit near Sapporo City in September In the protection bed #5, a 186 large part of the passage was filled by a sandbar because of the change in water flow 187 and current caused by the typhoon. 188 Experimental animals and transmitter attachment procedures 189 In the Toyohira River, adult chum and masu salmon migrate upstream from 190 September to October. All chum and masu salmon used in this study were captured 191 using a net km from the mouth of the Toyohira River and transferred to 11
13 192 outdoor tanks at the Chitose Salmon Aquarium for calibration of the EMG signals to 193 swimming speeds. In 2010, 3 males (fork length, cm; body weight kg) and 3 females (fork length, cm; body weight kg) chum salmon, 195 and 3 males (fork length, cm; body weight kg) and 3 females (fork 196 length, cm; body weight kg) masu salmon were studied. In 2011, male (fork length, 68.7 cm; body weight 3.3 kg) and 5 females (fork length, cm; body weight kg) chum salmon, and 2 males (fork length, cm; 199 body weight kg) and 1 female (fork length, 58.7 cm; body weight 2.6 kg) masu 200 salmon were studied. 201 Each fish was equipped with a cylindrical, epoxy-encased EMG transmitter 202 (CEMG-R11-35, Lotek Engineering Inc., Newmarket, Ontario, Canada; 18.3 g in air, mm in diameter, and 53.0 mm in length) attached externally to the body surface 204 and positioned in front of the dorsal fin using the procedure developed by Makiguchi et 205 al. (2011). External attachment is suitable for short-term telemetry research, which 206 reduces handling stress for the fish (Bridger and Booth 2003; Makiguchi et al. 2007). To 207 attach the tag, experimental fish were anaesthetized using FA100 (eugenol; Tanabe 208 Seiyaku Co. Ltd, Osaka, Japan) at a concentration of 0.5 ml l -1 in water from the 209 Chitose River and placed upright on a surgical table. Their gills were irrigated with 12
14 210 water containing diluted FA100 to maintain sedation during the attachment procedure. 211 Two stainless-steel needles large enough hold the restraining nylon ties were pushed 212 through the dorsal muscle to secure the EMG transmitter and sutured into place using 213 nylon ties and epoxy resin. Silicon pads were attached to minimize abrasion. The nylon 214 ties were passed through the needles and tied on the opposite side (Bridger and Booth ). The EMG transmitters consisted of an epoxy-coated transmitter package with a 216 pair of Teflon-coated electrodes with brass muscle-anchoring tips (dimension mm). The EMG electrodes were inserted subcutaneously using a hypodermic needle at a 218 ratio to the body length of approximately 0.7 on the left side of the fish. The electrodes 219 detected electrical potentials within the axial dark muscle, which contained red muscle 220 tissue. The amplitude and frequency of these pulses was directly correlated to the level 221 of muscle activity. Paired electrode tips were positioned approximately 1 cm apart and 222 secured in the lateral red muscle toward the rear of the fish. These muscles are primarily 223 used in steady, non-bursting, aerobic swimming activity (Beddow and McKinley 1999). 224 EMG signals are generally related to swimming speed (Økland et al. 1997). The 225 electrodes were sutured to avoid entangling vegetation or other environmental structures. 226 The CEMG model was equipped with a differential muscle probe, a signal conditioning 227 circuit, a digitizer, a microcontroller, and a radio transmitter. The voltage corresponding 13
15 228 to muscle activity was sampled at 2-s intervals. Individual samples were summed and 229 stored temporarily. At the end of the interval, the mean value was calculated, and an 230 activity level (EMG signal) ranging from 0 to 50 (no units) was assigned, and 231 transmitted to a radio receiver (model SRX_600; Lotek Engineering Inc.). The 232 attachment procedures usually required approximately 5 min to complete. 233 Calibration of EMG signals to swimming speeds 234 EMG signals were calibrated to swimming speed in the swim tunnel using 235 procedures developed by Makiguchi et al. (2011). The Chitose River water was poured 236 into the swim tunnel before each trial. The water temperature was set between 11.5 and ºC, which was similar to the water temperature at the capture point in the Toyohira 238 River. Experimental fish were individually placed into the swimming section of the 239 swim tunnel, and 10 EMG signals at 0 m s -1 were measured using the EMG radio 240 receiver when the fish maintained a holding position. Next, the trial was started at 0.3 m 241 s -1, and the water velocity was increased incrementally by 0.3 m s -1, with 10 EMG 242 signals measured at each increment. Fish readily swam against the current and rarely 243 came in contact with the grid at the back of the swim chamber. The trial ended at 1.2 m 244 s -1. We used the average EMG output at each velocity for each fish to calibrate 245 transmitter output to swimming speed. We evaluated the relationship between the mean 14
16 246 EMG output and water velocity using a linear regression analysis. 247 Field Study 248 Few studies report that fish with EMG transmitters were calibrated in respirometers 249 prior to their release. In September and October of 2010 and 2011, chum and masu 250 salmon used in the EMG-calibration experiment were individually released in the study 251 area and tracked on foot during upstream migration using a hand-held directional Yagi 252 antenna (Table 1). The position of each individual was determined using 253 EMG-transmitter signals received by three SRX_600 radio receivers. The radio 254 receivers recorded EMG signals at 2-s intervals. Migration time in each segment was 255 measured for each fish by subtracting the time on reaching the exit from the time at 256 entry. The chum and masu salmon were often observed to have stopped swimming 257 during their upstream migration, and more than 3 min of swimming cessation during 258 upstream migration was defined as holding behavior. The calculation of swimming time 259 did not include holding time. EMG signals were converted to swimming speed using the 260 equations established from the EMG calibration for each fish. After tracking, water 261 depth was measured at 0.2-m intervals in the water column, and water velocity was 262 measured at 2 5-m intervals along the tracking area. All tagged fish were released 263 below protection bed #5, and the tagged fish were tracked intermittently for 15
17 264 approximately 2 5 days. The number of times the fish exceeded Ucrit and the time spent 265 swimming over Ucrit were calculated from the time-course trajectory of swim speed, 266 using the Ucrit calculated in the laboratory for both species. 267 Statistical analyses 268 An analysis of covariance (ANCOVA) was used to compare the swimming abilities 269 of chum and masu salmon in the laboratory, and separate predictive relationships were 270 required for body length and body mass. ANCOVA was also used to compare the 271 swimming abilities of male and female in masu salmon in the laboratory. If the 272 ANCOVA was not be applicable in the statistical analysis, Student s t-test was used. 273 Statistical comparisons of the swimming performance of chum and masu salmon in 274 the field were analyzed using Student s t-test. The results were expressed as the mean ± 275 SE, and data were considered significant when P < For the relationship between swimming speed and MO2cor, regression analysis of 277 exponential equations using a parametric analysis of variance (ANOVA) were used. A 278 value of P < 0.05 was considered statistically significant Results 281 The statistical analysis by ANCOVA showed no significant parallelism of the 16
18 282 regression line for body length (P = > 0.05) or body mass (P = > 0.05) to 283 compare the swimming abilities of chum and masu salmon in the laboratory. In the test 284 of parallelism on regression line in the swimming performance (e.g. Ucrit) in the 285 laboratory study, Ucrit of male and female masu salmon was not significant (P = > ). Student s t-test also revealed no significant differences (P > 0.05) in the 287 swimming abilities in the laboratory or upstream-migration behavior in the field 288 between females and males of chum and masu salmon. 289 Swimming abilities in the laboratory 290 The Ucrit values for the chum and masu salmon were 1.42 BL s -1 and 1.89 BL s -1, 291 respectively, and masu salmon were significantly faster than chum salmon (P < 0.01; 292 Fig. 3). The chum and masu salmon SMRs were mgo2h 1 kg 1 and mgo2 293 h 1 kg 1, respectively, and the masu salmon s SMR was significantly lower than that of 294 chum salmon (P < 0.01; Fig. 3). The chum and masu salmon AMRs were mgo2 295 h 1 kg 1 and mgo2 h 1 kg 1, respectively. The chum and masu salmon AMSs were mgo2 h 1 kg 1 and mgo2 h 1 kg 1, respectively. The differences in AMR 297 and AMS values between the 2 salmon species were not significant (Fig. 3). 298 During the swimming test, the rate of oxygen consumption (MO2) tended to 299 increase proportionally with swimming speed. No increases in MO2 were observed at 17
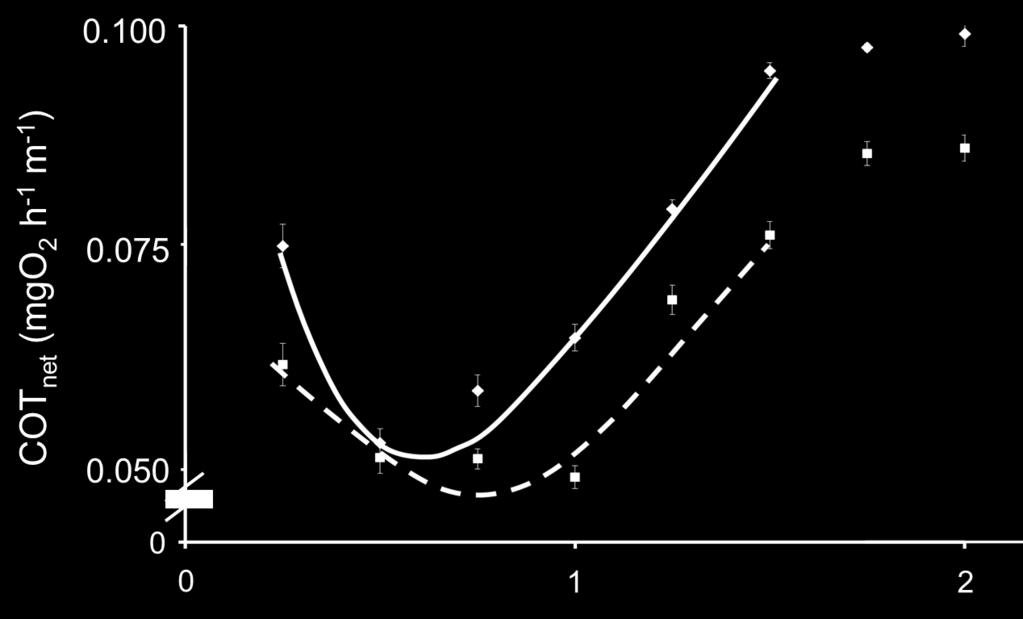
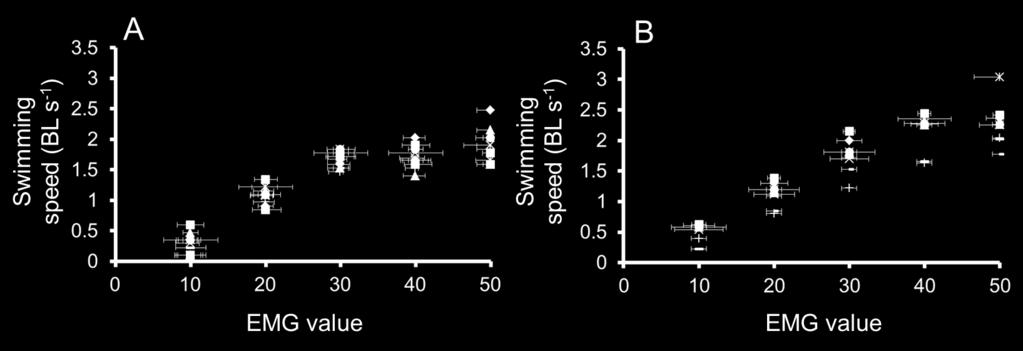
19 300 low swimming speeds in masu salmon (Fig. 4A). Figure 4B shows the relationship 301 between the MO2cor and swimming speed on an exponentially curve. The exponential 302 curves satisfactorily fitted the data for chum and masu salmon, and a significant 303 difference was found between chum and masu salmon (P < 0.05). The COTnet was also 304 influenced by swimming speed. It is worth noting that the lowest COTnet was found at and 1.0 BL s 1, which were considered the optimal swimming speeds in chum 306 salmon and masu salmon, respectively (Fig. 5). Although the COTnet of masu salmon 307 tended to be lower than for chum salmon, there was no significant difference between 308 the two species (P > 0.05). 309 From the EMG data, the muscle-activity levels were significantly correlated with 310 swimming speed according to simple regression analysis for all chum and masu salmon 311 (average of 12 chum salmon in 2010 and 2011, r 2 =0.92, P<0.05; average of 9 masu 312 salmon in 2010 and 2011, r 2 =0.97, P<0.05). A simple regression analysis was used to 313 convert EMG signals to swimming speeds in the field study by avoiding EMG values 314 above 40 showing no change in swimming speed. In the plots of all fishes, 315 U=0.043EMG for the chum salmon (Fig. 6A) and U=0.049EMG for 316 the masu salmon (Fig. 6B), where U is swimming speed (BL s -1 ) and EMG is the EMG 317 values. 18
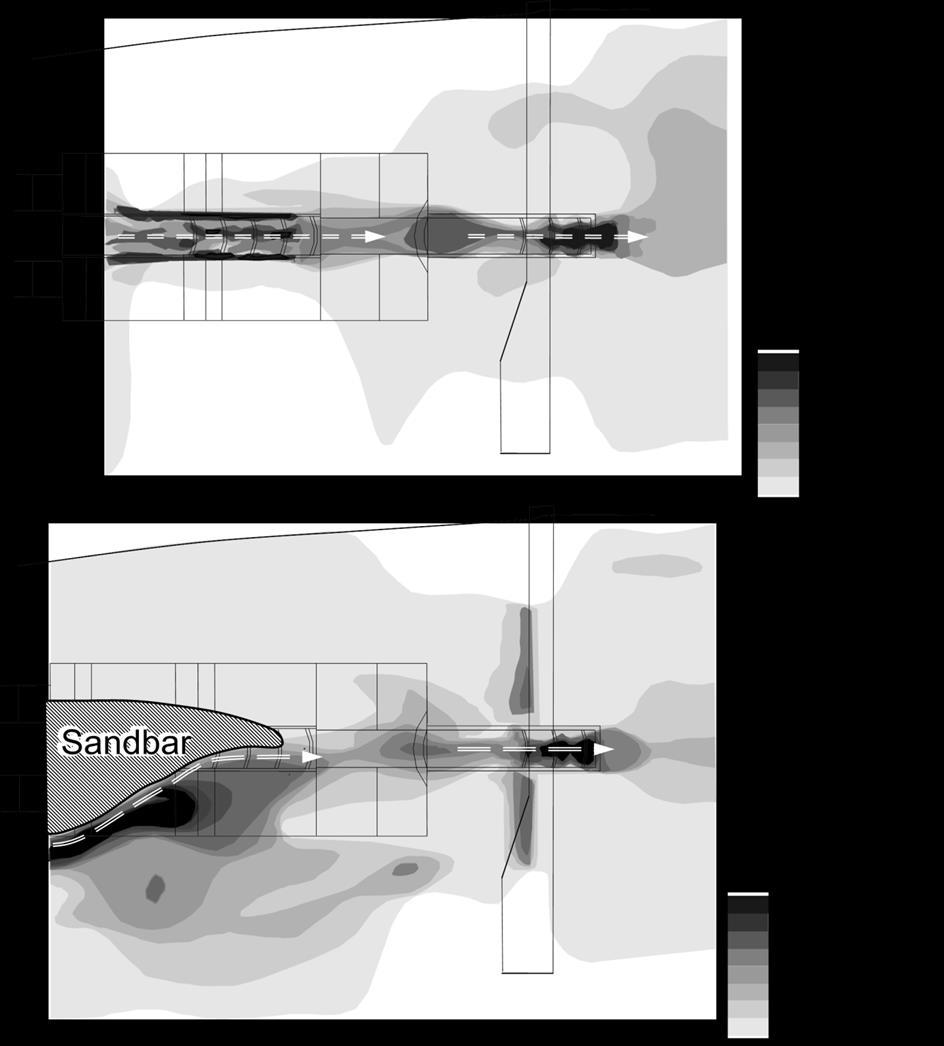
20 318 Upstream-migration behavior in the field 319 In 2010 and 2011, EMG data for all chum and masu salmon were collected around 320 the protection bed and groundsill #5. All tagged fish migrated upstream or downstream 321 using the fish passages on the day of release, and none of the fish that entered fish 322 passages at the protection bed and groundsill #5 made downstream migrations. Table shows the percentage of passing fish. The passage rate for both chum and masu salmon 324 at each structure dropped in At the fish passages of the ground sill #5, the passage 325 rate for masu salmon did not change, but only one fish was observed passing 326 successfully in Figures 7 shows the water velocity and passage path at the fish passages of the 328 protection bed and groundsill #5 in 2010 (A) and 2011 (B). In 2011, 60-80% of the fish 329 passage area at the protection bed #5was filled up by a sandbar because a severe 330 typhoon hit Sapporo City in September As a result, the water velocity was greater 331 in 2011 than in In 2010, all fish passed through the fish passages at the protection 332 bed and groundsill #5, but the fish only passed along the sandbar at the protection bed 333 # Table 2 shows the swimming speed and approach time at the fish passages of the 335 protection bed and groundsill #5 in 2010 and At the groundsill #5, chum and masu 19
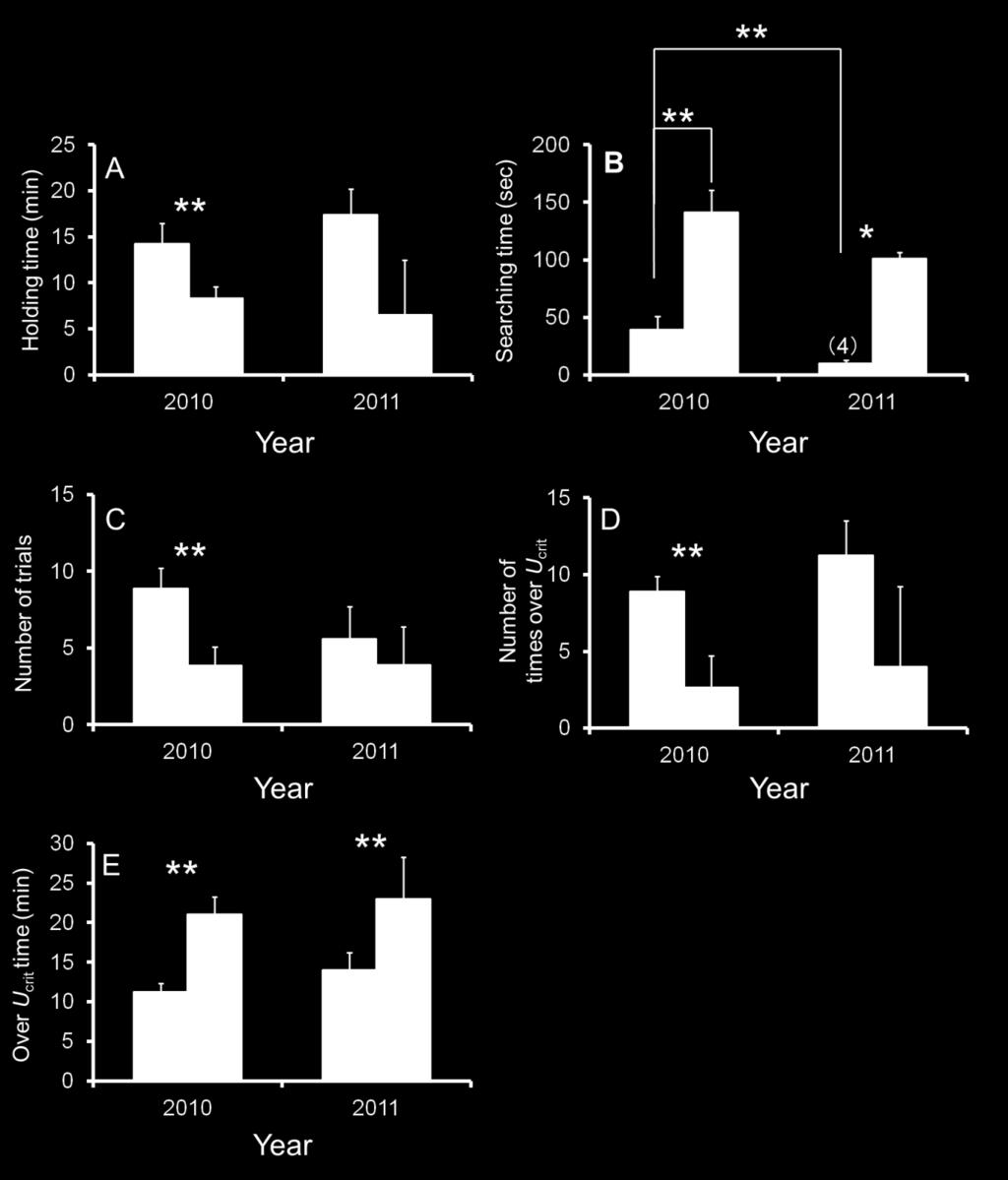
21 336 salmon showed no difference in swimming speed and approach time in either 2010 or Fig. 8 shows the time-course trajectory of swimming speeds of representative chum 339 salmon (A, C) and masu salmon (B, D) through the fish passages. Non-aerobic 340 swimming performance exceeding Ucrit was observed in both species at each step in the 341 fish passages. In the pools of the fish passages, each fish swam relatively slowly, 342 approaching the optimal swimming speed calculated in the laboratory. No significant 343 difference was found the approaching time between chum and masu salmon at each step 344 (P > 0.05). 345 The holding time at each step through the fish passage at the protection bed and 346 groundsill #5 was significantly shorter for masu salmon than chum salmon in 2010 (P < ; Fig. 9A), and the search time at each step through the fish passage at the 348 protection bed and groundsill #5 was significantly longer for masu salmon than chum 349 salmon in 2010 (P < 0.01) and 2011 (P < 0.05; Fig. 9B). There was also a significant 350 difference in the search time for chum salmon between 2010 and 2011 (P < 0.01; Fig B). Fig. 9C shows the number of trial times for each step through fish passage at the 352 protection bed and groundsill #5. The number of trial times for the protection bed and 353 groundsill #5 in 2010 was significantly shorter for masu salmon than chum salmon (P < 20
22 ). The number of times that fish were observed swimming at speeds greater than 355 Ucrit at the protection bed and groundsill #5 was significantly lower for masu salmon 356 than chum salmon in 2010 (Fig. 9D). In addition, the time over Ucrit for the protection 357 bed and groundsill #5 was significantly longer for masu salmon than chum salmon in 358 both 2010 and 2011 (P < 0.01; Fig. 9E) Discussion 361 Swimming ability, measured using a swim tunnel in the laboratory, and upstream 362 spawning-migration behavior, measured using EMG telemetry at the fish passages in 363 the Toyohira River, were compared between chum and masu salmon. Masu salmon are 364 better adapted to swimming in a high-water-velocity area for an extended periods of 365 time than chum salmon, and this adaptability could make masu salmon more successful 366 than chum salmon in completing a migration to their spawning ground in the upper 367 reaches of the river. 368 Both the mean MO2 throughout the swimming test and the COTnet tended to be 369 lower in masu salmon than chum salmon. This pattern was particularly obvious above BL s 1, where the COTnet increased with speed in a parallel in the two species until 2 BL 371 s 1 (Fig. 5). Excess post-exercise oxygen consumption (EPOC) is estimated as the time 21
23 372 required to return to their normal level of oxygen consumption (recovery time) and the 373 total oxygen cost of swimming to Ucrit (Brett 1964; Lee et al. 2003b). Lee et al. (2003b) 374 and Farrell (2007) suggested that increase in the COT rate was estimated % lower 375 than it was without EPOC. Although we did not examine EPOC in this study, the 376 COTnet increased in parallel with increasing the swimming speed over 1 BL -1 s in chum 377 and masu salmon. 378 Lee et al. (2003a) also showed that the MO2 routine, with properties similar to the 379 SMR, was no different between coho and sockeye salmon captured around the same 380 time. Both Ucrit and the optimal swimming speed were significantly faster for masu 381 salmon than chum salmon, and the SMR of masu salmon was significantly lower than 382 that of chum salmon in this study. Masu salmon spawning grounds are usually located 383 in the upper reaches of rivers (Kato 1991). Therefore, it is likely that masu salmon must 384 pass through more cross-river constructs with high-water-velocity than chum salmon. In 385 high-water-flow areas, masu salmon may swim use aerobic swimming more 386 successfully than chum salmon. As masu salmon stay in the river longer time than chum 387 salmon, the metabolic rate of masu salmon is lower than chum salmon. Therefore, it 388 was reasonable to assume that the basal metabolism of the masu salmon was lower than 389 chum salmon, and there was difference in slope of MO2cor between chum and masu 22
24 390 salmon. Lee et al. (2003a) showed that salmons, migrating for long-distance, the 391 attained a significantly higher Ucrit at and were more efficient swimmers at Ucrit because 392 of a lower MO2max compared with other salmons migrating for a comparatively short 393 distance. Masu salmon have higher Ucrit and lower SMR than chum salmon. However, 394 AMR was similar between chum and masu salmon in this study presumably because 395 that small sample sizes may have prevented detecting differences. 396 It is well known that water temperature influences the metabolism and swimming 397 performance of salmonids (Lee et al. 2003a; Farrell 2007). This study was carried out at 398 the water temperature of the Chitose River in autumn. It should be mentioned that a 399 significant change might be observed in the metabolism and swimming performance of 400 masu salmon at a higher water temperature (as in summer, before masu salmon start 401 their upstream migration). Lee et al. (2003a) investigated relationships between 402 swimming performance and metabolism for sockeye salmon of different stocks in the 403 swim test and found that fishes captured in the upper reaches of the river migrates much 404 longer distances and negotiate more severe hydraulic challenges than the coastal fish. 405 Compared to chum salmon, masu salmon have their spawning grounds farther upriver 406 and remain in the river longer time; therefore, masu salmon are better adapted to swim 407 for extended time in an area of high-water-velocity. 23
25 408 The MO2 of both species tended to plateau before reaching Ucrit (Fig.4A), and the 409 slopes of the COTnet shifted between 1.75 BL s -1 and 2 BL s -1 in both species (Fig.5), the 410 slopes of the EMG shifted between 1.5 BL s -1 and 2 BL s -1 in both species (Fig. 6). The 411 amplitude and frequency of these pulses was directly correlated to the level of 412 red-muscle activity (Beddow and McKinley 1999; Økland et al. 1997), indicating that 413 the shift in muscle performance from white muscle to red muscle occurred between BL s -1 and 2 BL s -1 ; this result suggests that supplementary swimming effort when 415 approaching Ucrit was sustained by anaerobic metabolism. Farrell (2007) indicated that 416 the oxygen requirements should either plateau or decrease at nearly Ucrit because of the 417 activation of white muscle (based on electro-myography recordings) and glycolysis 418 (based on glycogen depletion and lactate accumulation in white muscle); however, 419 Farrell (2007) also explained that white-muscle contraction has a significant aerobic 420 component that drives an increase in MO2 near Ucrit. White muscle has aerobic capacity 421 and is activated at speeds below Ucrit. Therefore, the metabolic cost of swimming 422 increases exponentially with velocity (Fig. 4B). 423 In the field study, the searching time was significantly longer for masu salmon than 424 chum salmon, and non-aerobic swimming performance exceeding Ucrit was necessary 425 for both two species at each step in the fish passages because they had to swim at high 24
26 426 flow rate (exceeding 2m -1 s). It is likely that masu salmon could search long periods of 427 time in the each pool in order to find easy routes for the reduction of energy loss. 428 Gowans et al. (1999) reported that migrating Atlantic salmon (Salmo salar) sometimes 429 failed to locate the entrance to a pool of fish passage and appeared to be attracted by 430 tailrace and turbine flows. Makiguchi et al. (2011) indicated that it was important for the 431 successful migration that salmon find passage upstream rapidly and easily without 432 wandering. Our results suggested that masu salmon search approach routes, reducing 433 energy expended in approaching the fish passages compared to chum salmon. 434 Hinch and Bratty (2000) tracked adult sockeye salmon during upstream migration 435 using EMG telemetry at Hell s Gate in the Fraser River and reported that swimming 436 speed patterns alternated at different time scales between relatively fast and slow speeds. 437 In our study, the search time was significantly longer for masu salmon than chum 438 salmon, but the number of times over Ucrit at each fish passages were significantly lower 439 for masu salmon than chum salmon. Masu salmon could select slow and rapid 440 swimming, depending on the river obstacle, and chum and masu salmon searched for 441 routes of approach at speeds approximating the optimal swimming speed. Hinch and 442 Rand (2000) reported that successful sockeye salmons swan at the Hell s Gate, and 443 migrated according to an optimal swimming speed model range in whole of Fraser 25
27 444 River, which migrants minimized transport costs per unit distance. Our study also 445 showed that successful chum and masu salmon used the energy-saving swimming at 446 fish passages. In fact the fish passages were organized by some steps to secure 447 recovering time in the Toyohira River, successful chum and masu salmon could almost 448 use optimal swimming speed except over the Ucirt swimming at each step. 449 Chum salmon, however, had longer holding times and shorter search times than 450 masu salmon at each fish passage. Because the holding time occurred when each fish 451 crossed to the next step in the fish passage, this finding indicated that the holding time 452 occurred before or after the burst speed over Ucrit. During upstream migration, chum 453 salmon had a consistent holding behavior after swimming at high speed over Ucrit, 454 indicating that they swim at speeds greater than Ucrit prior to holding (Makiguchi et al., ). Chum salmon showed the more trial than masu salmon at each fish passage, 456 indicating that chum salmon swim over Ucrit. 457 Lee et al. (2003b) reported that routine oxygen consumption was restored within 458 approximately 1 h in adult sockeye salmon in the EPOC test. In our study, if the holding 459 time was defined as the recovery time after excess swimming, chum salmon spent 460 approximately minutes in recovery at each steps. These observations may show 461 that chum salmon suffered an energy loss at the fish passages; extended periods of 26
28 462 swimming at fast speeds may deplete the limited energy reserves of fish and leave little 463 energy for additional upriver progress (Rand and Hinch 1998). It is also possible that 464 the sustained and repeated bouts of anaerobic activity (e.g., swimming at speeds over 465 Ucrit for prolonged periods of time) could lead to mortality by causing lethal levels of 466 blood lactate to accumulate, resulting in metabolic acidosis (Hinch and Bratty, 2000). It 467 is possible that the loss of stored energy decreased the passing rate of chum salmon at 468 the groundsill #5 (see Table 2). Moreover, swimming over Ucrit caused the fish fatigue 469 and stress. Webb (1971) reported that anaerobic metabolism occurred during swimming 470 at 80% of Ucrit. Chum salmon were considered more stressed than masu salmon because 471 the Ucrit of chum salmon were lower than that of masu salmon. 472 In contrast, masu salmon required only approximately 10 minutes of recovery time, 473 suggesting two possibilities. First, masu salmon may be resilient to exceeding Ucrit. 474 Second, no more than 10 minutes was required to pass though the protection bed and 475 groundsill #5 in the Toyohira River. For example, the total length of the fish passage of 476 the protection bed and groundsill #5 was less than 50 m (8 pools; see Fig. 2), but the 477 vertical-slot fishway at the Seton River and the Fraser River, British Columbia, is more 478 than100 m long (Hinch et al. 1996; Pon et al. 2009a). Nevertheless, more information is 479 needed regarding energy cost and metabolism (e.g., EPOC) and spawning behavior in 27
29 480 masu and chum salmon. 481 Percentages of fish passing the fish passage of the protection bed #5 were 67 % and % in chum and masu salmon, respectively. It was unclear if the differences between 483 successful and unsuccessful passages were due to swimming speed and approach time; 484 however, it was likely that unsuccessful individuals gave up quickly at the fish passages 485 or required a very long time to find passing routes. No significant difference was 486 observed between 2010 and 2011 in migration behavior at each fish passage, except in 487 the search time of chum salmon. The mean search time of masu salmon was lower in than 2010, when the routes had been filled up by a sandbar during the typhoon. 489 In conclusion, comparisons between chum salmon and masu salmon of both 490 swimming ability in the laboratory swim tunnel and upstream-migration behavior using 491 EMG telemetry at fish passages in the Toyohira River provided strong evidence that 492 masu salmon were better adapted than chum salmon to swimming for an extended 493 periods of time in areas of high-water-velocity. Masu salmon were better able to migrate 494 upstream to their spawning grounds in the upper reaches of the river than chum salmon. 495 In this study, swimming performance did not vary significantly with body length or 496 mass; however, these differences should be analyzed by further morphological and 497 genetic studies. 28
30 Acknowledgements 500 We thank the following people and organizations for their help and cooperation: Mr. 501 M. Kikuchi of the Chitose Salmon Aquarium for the use of the Chitose Salmon 502 Aquarium facilities, the staffs of the Sapporo Salmon Museum for valuable information 503 about the Toyohira River, and the Sapporo River Office, Hokkaido Regional 504 Development Bureau for valuable input on the selection of the experimental site References 507 Beddow, T.A., and McKinley, R.S Importance of electrode positioning in 508 biotelemetry studies estimating muscle activity in fish. J. Fish Biol. 54(4): doi: /j tb02035.x 510 Bell, W.M. and Terhune, L.D.B Water tunnel design for fisheries research. Tech. 511 Rep. Fish. Res. Bd Can. 195: Brett, J.R The respiratory metabolism and swimming performance of young 513 sockeye salmon. J. Fish. Res. Bd Can. 21(5): doi: /f Bridger, C.J., and Booth, R.K The effects of biotelemetry transmitter presence 515 and attachment procedures on fish physiology and behavior. Rev. Fish. Sci. 11(1): 29
31 Claireaux, G., Couturier, C., and Groison, A.L Effect of temperature on 518 maximum swimming speed and cost of transport in juvenile European sea bass 519 (Dicentrarchus labrax). J. Exp. Biol. 209(17): doi: /jeb Farrell, A.P Cardiorespiratory performance during prolonged swimming tests 521 with salmonids: a perspective on temperature effects and potential analytical pitfalls. 522 Phil. Trans. R. So. B. 362(1487): doi: /rstb Fry, F. E. J. (1971). The effect of environmental factors on the physiology of fish. In 524 Fish Physiology. Edited by W.S. Hoar, and D. J. Randall. Academic Press, New 525 York, NY. pp Gamperl, A. K., Rodnick, K. J., Faust, H. A., Venn, E. C., Byington, J. D., Bennett, M. 527 T., Crawshaw, L. I., Kelley, E. R., Powell, M. S. and Li, H Metabolism, 528 swimming performance and tissue biochemistry of high desert redband trout 529 (Oncorhynchus mykiss ssp.): Evidence of phenotypic differences in physiological 530 function. Physiol. Biochem. Zool. 75(5): Gehrke, P.C., Fidler, L.E., Mense, D.C., and Randall, D.J A respirometer with 532 controlled water-quality and computerized data acquisition for experiments with 533 swimming fish. Fish Physiol. Biochem. 8(1): doi: /BF
32 534 Gowans, A. R. D., J. D Armstrong and I. G. Priede, Movements of adult Atlantic 535 salmon in relation to a hydroelectric dam and fish ladder. J Fish Biol. 54(6): doi: /j tb02028.x 537 Hinch, S.G., Diewert, R.E., Lissimore, T.J., Prince, A.M.J., Healey, M.C., and 538 Henderson, M.A Use of electromyogram telemetry to assess difficult 539 passage areas for river-migrating adult sockeye salmon. Trans. Am. Fish. Soc (2): doi: / (1996)125<0253:UOETTA>2.3.CO; Hinch, S.G., and Bratty, J Effects of swim speed and activity pattern on success 542 of adult sockeye salmon migration through an area of difficult passage. Trans. Am. 543 Fish. Soc. 129(2): doi: / (2000)129<0598:EOSSAA>2.0.CO; Hinch, S.G., and Rand, P.S Optimal swimming speeds and forward-assisted 546 propulsion: energy-conserving behaviours of upriver-migrating adult salmon. Can. 547 J. Fish. Aquat. Sci. 57: doi: /f Kato, F Life history of masu and amago salmon (Oncorhynchus masou and 549 Oncorhynchus rhodurus). In Pacific Salmon Life Histories. Edited by C. Groot and 550 L. Margolis. University of British Columbia Press, Vancouver, B.C. pp Lee, C.G., Farrell, A.P., Lotto, A., MacNutt, M.J., Hinch, S.G., and Healey, M.C. 2003a. 31
33 552 The effect of temperature on swimming performance and oxygen consumption in 553 adult sockeye (Oncorhynchus nerka) and coho (O. kisutch) salmon stocks. J. Exp. 554 Biol. 206(18): doi: /jeb Lee, C.G., Farrell, A.P., Lotto, A., Hinch, S.G., and Healey, M.C. 2003b. Excess 556 post-exercise oxygen consumption in adult sockeye (Oncorhynchus nerka) and 557 coho (O. kisutch) salmon following critical speed swimming. J. Exp. Biol. 206(18): doi: /jeb Luna-Acosta, A., Lefrançois, C., Millot, S., Chatain, B., and Bégout, M.L Physiological response in different strains of sea bass (Dicentrarchus labrax): 561 Swimming and aerobic metabolic capacities. Aquaculture 317(1-4): doi: /j.aquaculture Makiguchi, Y., Nii, H., Nakao, K., and Ueda, H Upstream migration of adult 564 chum and pink salmon in the Shibetsu River. Hydrobiologia 582: doi: /s Makiguchi, Y., Konno, Y., Konishi, K., Miyoshi, K., Sakashita, T., Nii, H., Nakao, K., 567 and Ueda, H EMG telemetry studies on upstream migration of chum salmon 568 in the Toyohira River, Hokkaido, Japan. Fish Physiol. Biochem. 37(2): doi: /s y. 32
34 570 Mayama, H Studies on the freshwater life and propagation technology of masu 571 salmon, Oncorhynchus masou (Brevoort). Sci. Rep. Hokkaido Salmon Hatch. 46: (in Japanese with English summary). 573 Økland, F., Finstad, B., McKinley, R.S., Thorstad, E.B., and Booth, R.K Radio-transmitted electromyogram signals as indicators of physical activity in 575 Atlantic salmon. J. Fish Biol. 51(3): doi: /j tb01505.x. 577 Pon, L. B., Cooke, S. J. and Hinch, S. G Passage efficiency and migration 578 behaviour of salmonid fishes at the Seton Dam fish passage. Final report for the 579 Bridge Coastal Restoration Program. Project 05.Se Pon, L.B., Hinch, S.G., Cooke, S.J., Patterson, D.A., and Farrell, A.P. 2009a. A 581 Comparison of the physiological condition, and fish passage time and success of 582 migrant adult sockeye salmon at Seton River Dam, British Columbia, under three 583 operational water discharge rates. N. Am. J. Fish. Manage. 29(5): doi: /M Pon, L.B., Hinch, S.G., Cooke, S.J., Patterson, D.A., and Farrell, A.P. 2009b. 586 Physiological, energetic and behavioural correlates of successful fish passage of 587 adult sockeye salmon Oncorhynchus nerka in the Seton River, British Columbia. J 33
35 588 Fish Biol. 74(6): doi: /j x. 589 Rand, P. S., and S. G. Hinch Swim speeds and energy use of up-river migrating 590 sockeye salmon: simulating metabolic power and assessing risk of energy depletion. 591 Can. J. Fish. Aquat. Sci 55(8): doi: /f Roscoe, D.W., and Hinch, S.G Effectiveness monitoring of fish passage facilities: 593 historical trends, geographic patterns and future directions. Fish Fish. 11(1): doi: /j x. 595 Salo, E. O. (1991). Life history of chum salmon (Oncorhynchus keta). In Pacific Salmon 596 Life Histories. Edited by C. Groot and L. Margolis. University of British Columbia 597 Press, Vancouver, B.C. pp Schurmann, H., and Steffensen, J.F Effects of temperature, hypoxia and activity 599 on the metabolism of juvenile Atlantic cod. J. Fish Biol. 50(6): doi: /j tb01645.x. 601 Webb, P.W Swimming Energetics of Trout.2. Oxygen Consumption and 602 Swimming Efficiency. J. Exp. Biol. 55(2): Wagner, G. N., Kuchel, L. J., Lotto, A., Patterson, D. A., Shrimpton, M., Hinch, S. G., 604 and Farrell, A. P Routine and Active Metabolic Rates of Migrating Adult 605 Wild Sockeye Salmon (Oncorhynchus nerka Walbaum) in Seawater and Freshwater. 34
36 606 Physiol. Biochem. Zool. 79(1):
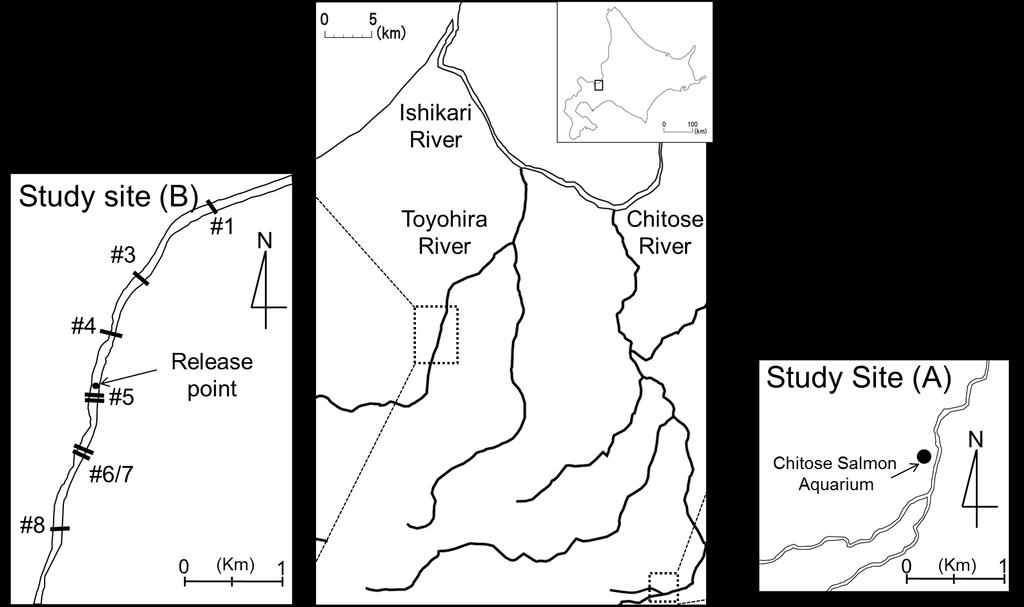
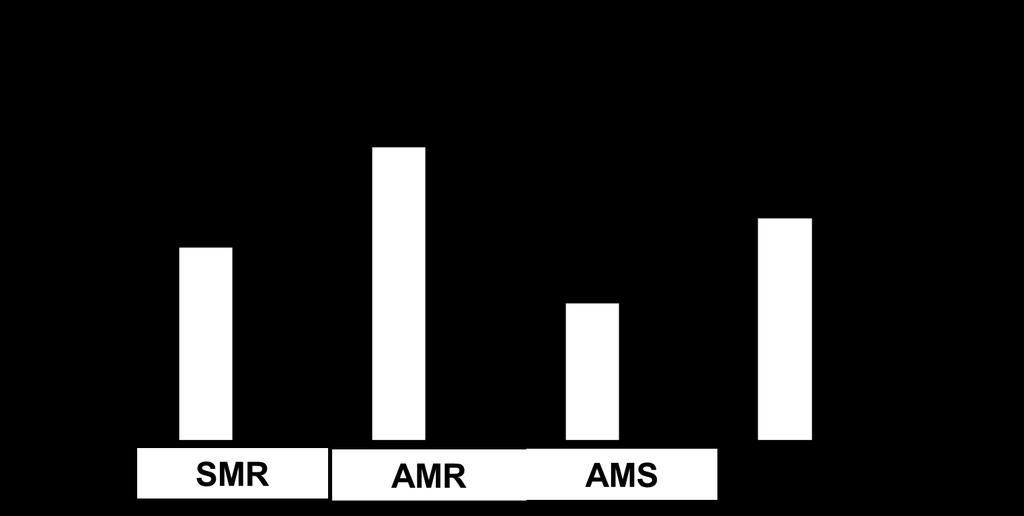
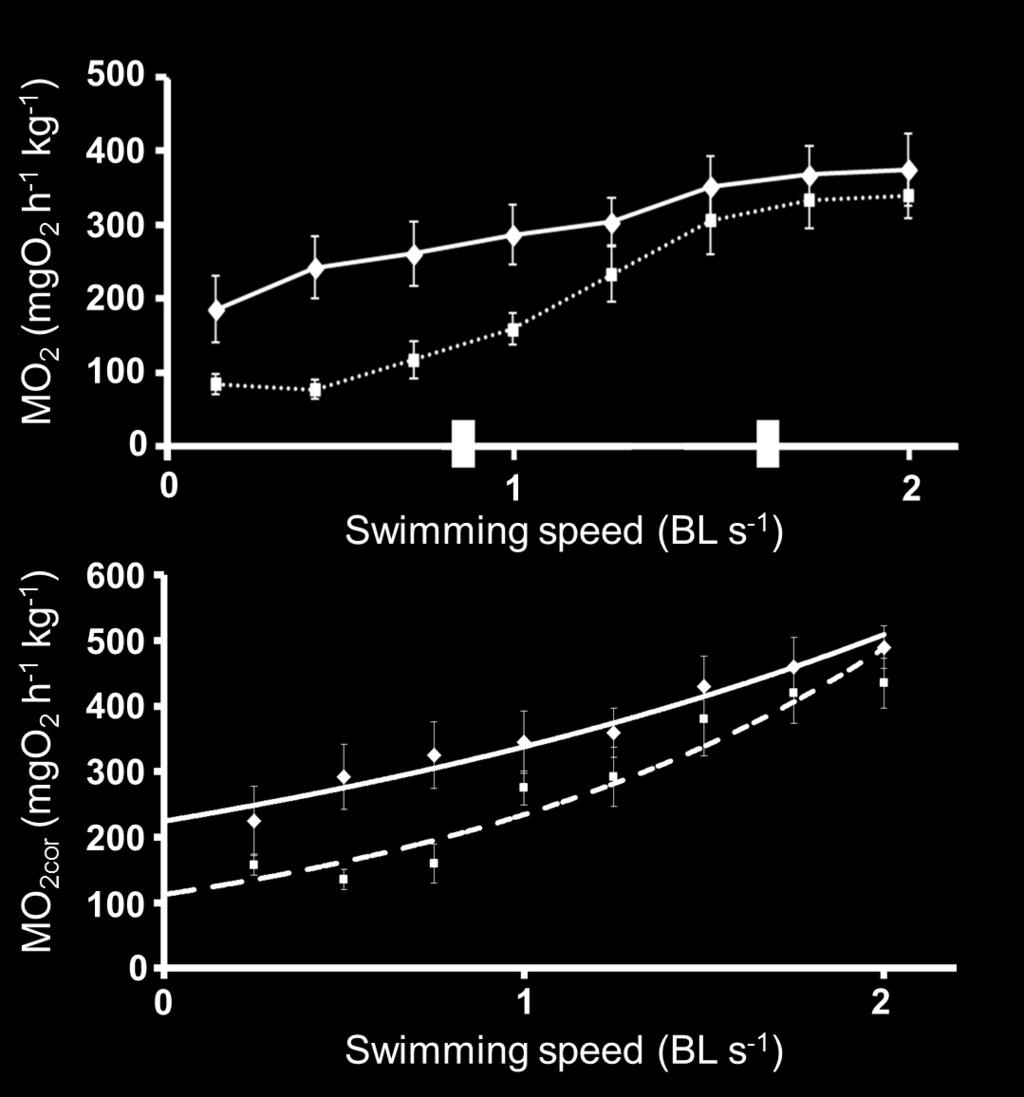
37 Figure Captions Figure 1. A map of the study site showing the experimental site in the Chitose Salmon Aquarium near the Chitose River (A), the positions of the groundsills, and chum and masu salmon release point in the Toyohira River (B) Figure 2. A diagram of the fish passage of the protection bed #5 (a) and fish passage of the groundsill #5 (b). The dotted line represents the water s edge for a typical water level in The number of circles in the fish passage is the pool number. Figure 3. Average (± SE) standard metabolic rate (SMR), active metabolic rate (AMR), aerobic metabolic scope (AMS), and critical swimming speed (Ucrit) for chum salmon (white column) and masu salmon (black column). The left Y axis corresponds to oxygen consumption (MO2 in mgo2 h 1 kg 1 ) related to SMR, AMR, and AMS. The right Y axis corresponds to swimming speed (BL s 1 ) for Ucrit. The asterisk indicates significant differences by Student s t-test (P < 0.01). Figure 4. (A) The relationship between the rate of oxygen consumption (MO2) and swimming speed (BL s 1 ) for chum salmon (solid line) and masu salmon (dotted line). (B) The relationship between the MO2cor and swimming speed (BL s 1 ) for chum salmon (solid line) and masu salmon (dotted line). The relationship is exponential; for chum salmon, MO2cor = e0.409u (P < 0.05; r 2 = 0.952); for 36
38 masu salmon, MO2cor = e0.734u (P < 0.05; r 2 = 0.903). Figure 5. The relationships between net cost of transport (COTnet) and swimming speed (BL s 1 ) for chum salmon (solid line) and masu salmon (dotted line). The COTnet varied for chum salmon according to the polynomial equation COTnet= 3.55 e e U 2 - U (r 2 =0.911) and for masu salmon according to the polynomial equation COTnet= 3.91 e e U 2 - U (r 2 =0.920). Figure 6. The relationships between electromyogram (EMG) signals and swimming speed (BL s -1 ) for chum salmon (A: n=12) and masu salmon (B: n=9) in all fish in the field study in 2010 and The standard error of each EMG value is expressed. Figure 7. The swimming path (red dotted line) of passing chum and masu salmon and water velocity at the fish passage for the protection bed and groundsill #5 in 2010 (A) and 2011(B). Figure 8. Time-series plots of swimming speeds of representative chum salmon in 2010(A) and 2011(C), and masu salmon in 2010 (B) and 2011(D). The dotted line represents the mean critical swimming speed (Ucrit; 1.42 BL -1 s or 1.89 BL -1 s) and optimal swimming speed (Uopt; 0.5 BL -1 s or 1.0 BL -1 s) in chum salmon and masu salmon, respectively. The number of circles in the fish passage is the pool number 37
39 in Fig. 2. Figure 9. The holding time (A), searching time (B), number of trials (C), number of times over Ucrit (D) and over Ucrit time (E) at each fish passage step in the fish passage of the protection bed and groundsill #5 for chum salmon (white column) and masu salmon (black column) in 2010 and The asterisk indicates significant differences by Student s t-test (P < 0.01, 0.05). The number of fish is provided in parentheses. 38
40 Table. 1 Year Species Releasing point Releasing date Number of experimental fish (female) Number of upstream migrating fish (female) Number of passing fish (female) Percentage of passing fish at the fishway #5 protection bed #5 ground sill #5 protection bed #5 ground sill #5 protection bed #5 ground sill 2010 chum 2010/10/6 6(3) 4(2) 4(2) 4(2) 2(2) #5 masu 2010/9/26 6(3) 6(3) 6(3) 6(3) 6(3) protection 2011 chum 2011/10/4 6(5) 6(5) 4(3) 4(3) bed masu 2011/9/25 3(1) 3(1) * Data in parentheses are the number of female fish
41 Table. 2 Year Species Swimming speed (BL/s)* Approaching time (min)* #5 Protection #5 Protection #5 ground sill bed bed #5 ground sill 2010 chum 0.87 ± 7.3 (4) 1.33 (2) 38.2 ± 28.0 (4) 45.1 (2) masu 1.13 ± 6.6 (6) 1.28 ± 6.5 (6) 25.3 ± 19.2 (6) 58.9 ±37.1 (6) 2011 chum 1.14 ± 2.6 (4) 1.34 (1) 25.3 ± 16.5 (4) 16.5 (1) masu 1.38 ± 4.3 (3) 1.24 (1) 37.2 ± 4.3 (3) 16.0 (1) * Values represent the mean ± standard error ** Data in parentheses are the number of female

42 Fig. 1

43 Fig. 2

44 Fig. 3
45 Fig. 4
46 Fig. 5
47 Fig. 6

48 Fig.7


49 Fig.8
50 Fig.9
Migratory behaviour of adult chum salmon, Oncorhynchus keta, in a reconstructed reach of the Shibetsu River, Japan
 Fisheries Management and Ecology Fisheries Management and Ecology, 28, 15, 425 433 Migratory behaviour of adult chum salmon, Oncorhynchus keta, in a reconstructed reach of the Shibetsu River, Japan Y.
Fisheries Management and Ecology Fisheries Management and Ecology, 28, 15, 425 433 Migratory behaviour of adult chum salmon, Oncorhynchus keta, in a reconstructed reach of the Shibetsu River, Japan Y.
Follow this and additional works at:
 University of Massachusetts - Amherst ScholarWorks@UMass Amherst International Conference on Engineering and Ecohydrology for Fish Passage International Conference on Engineering and Ecohydrology for Fish
University of Massachusetts - Amherst ScholarWorks@UMass Amherst International Conference on Engineering and Ecohydrology for Fish Passage International Conference on Engineering and Ecohydrology for Fish
Upstream migration of adult chum and pink salmon in. The original publication is available at Instructions for use
 Title Upstream migration of adult chum and pink salmon in Author(s)Makiguchi, Y.; Nii, H.; Nakao, K.; Ueda, H. CitationHydrobiologia, 582(1): 43-54 Issue Date 2007-05 Doc URL http://hdl.handle.net/2115/22080
Title Upstream migration of adult chum and pink salmon in Author(s)Makiguchi, Y.; Nii, H.; Nakao, K.; Ueda, H. CitationHydrobiologia, 582(1): 43-54 Issue Date 2007-05 Doc URL http://hdl.handle.net/2115/22080
Effects of Swim Speed and Activity Pattern on Success of Adult Sockeye Salmon Migration through an Area of Difficult Passage
 Transactions of the American Fisheries Society 129:598 606, 2000 Copyright by the American Fisheries Society 2000 Effects of Swim Speed and Activity Pattern on Success of Adult Sockeye Salmon Migration
Transactions of the American Fisheries Society 129:598 606, 2000 Copyright by the American Fisheries Society 2000 Effects of Swim Speed and Activity Pattern on Success of Adult Sockeye Salmon Migration
Sex differences in metabolic rate and swimming performance in pink salmon (Oncorhynchus gorbuscha): the effect of male secondary sexual traits
: the effect of male secondary sexual traits") Ecology of Freshwater Fish 2016 Ó 2016 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd ECOLOGY OF FRESHWATER FISH Sex differences in metabolic rate and swimming performance in pink salmon (Oncorhynchus
Ecology of Freshwater Fish 2016 Ó 2016 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd ECOLOGY OF FRESHWATER FISH Sex differences in metabolic rate and swimming performance in pink salmon (Oncorhynchus
Scott G. Hinch 1,2,, Emily M. Standen 1, Michael C. Healey 2,3 & Anthony P. Farrell 4
 Hydrobiologia 483: 147 160, 2002. E.B. Thorstad, I.A. Fleming & T.F. Næsje (eds), Aquatic Telemetry. 2002 Kluwer Academic Publishers. Printed in the Netherlands. 147 Swimming patterns and behaviour of
Hydrobiologia 483: 147 160, 2002. E.B. Thorstad, I.A. Fleming & T.F. Næsje (eds), Aquatic Telemetry. 2002 Kluwer Academic Publishers. Printed in the Netherlands. 147 Swimming patterns and behaviour of
Trout behavior and responses to pulsed flows: investigations utilizing electromyogram telemetery
 Trout behavior and responses to pulsed flows: investigations utilizing electromyogram telemetery Sarah A. Hamilton, Dennis E. Cocherell, Gardner J. Jones, and Javier B. Miranda, A. Peter Klimley, Lisa
Trout behavior and responses to pulsed flows: investigations utilizing electromyogram telemetery Sarah A. Hamilton, Dennis E. Cocherell, Gardner J. Jones, and Javier B. Miranda, A. Peter Klimley, Lisa
For next Thurs: Jackson et al Historical overfishing and the recent collapse of coastal ecosystems. Science 293:
 For next Thurs: Jackson et al. 2001. Historical overfishing and the recent collapse of coastal ecosystems. Science 293: 629-637. Resource: means of supplying a want or deficiency, from French resourdre
For next Thurs: Jackson et al. 2001. Historical overfishing and the recent collapse of coastal ecosystems. Science 293: 629-637. Resource: means of supplying a want or deficiency, from French resourdre
Excess post-exercise oxygen consumption in adult sockeye (Oncorhynchus nerka) and coho (O. kisutch) salmon following critical speed swimming
 and coho (O. kisutch) salmon following critical speed swimming") The Journal of Experimental Biology 26, 3253-326 23 The Company of Biologists Ltd doi:11242/jeb548 3253 Excess post-exercise oxygen consumption in adult sockeye (Oncorhynchus nerka) and coho (O kisutch)
The Journal of Experimental Biology 26, 3253-326 23 The Company of Biologists Ltd doi:11242/jeb548 3253 Excess post-exercise oxygen consumption in adult sockeye (Oncorhynchus nerka) and coho (O kisutch)
Technical Report 99-4 EFFECTS OF A SHAD FISHERY ON PASSAGE OF ADULT CHINOOK SALMON THROUGH THE OREGON-SHORE FISHWAY LADDER AT THE DALLES DAM
 Technical Report 99-4 EFFECTS OF A SHAD FISHERY ON PASSAGE OF ADULT CHINOOK SALMON THROUGH THE OREGON-SHORE FISHWAY LADDER AT THE DALLES DAM - 1996 A report for Project MPE-P-95-1 by C.A. Peery, T.C. Bjornn,
Technical Report 99-4 EFFECTS OF A SHAD FISHERY ON PASSAGE OF ADULT CHINOOK SALMON THROUGH THE OREGON-SHORE FISHWAY LADDER AT THE DALLES DAM - 1996 A report for Project MPE-P-95-1 by C.A. Peery, T.C. Bjornn,
INFLUENCE OF WATER TEMPERATURE ON ADULT SALMON AND STEELHEAD PASSAGE AND BEHAVIOR AT LOWER GRANITE DAM, 2008
 Technical Report 2009-7 INFLUENCE OF WATER TEMPERATURE ON ADULT SALMON AND STEELHEAD PASSAGE AND BEHAVIOR AT LOWER GRANITE DAM, 2008 by T.S. Clabough, M.A. Jepson, C.C. Caudill, and C.A. Peery 1 Department
Technical Report 2009-7 INFLUENCE OF WATER TEMPERATURE ON ADULT SALMON AND STEELHEAD PASSAGE AND BEHAVIOR AT LOWER GRANITE DAM, 2008 by T.S. Clabough, M.A. Jepson, C.C. Caudill, and C.A. Peery 1 Department
*Present address: University of Durham, Department of Biological Sciences, South Road, Durham DH1 3LE, UK.
 J. exp. Biol. 175, 325 331 (1993) Printed in Great Britain The Company of Biologists Limited 1993 325 SHORT COMMUNICATION AN ANNULAR RESPIROMETER FOR MEASURING AEROBIC METABOLIC RATES OF LARGE, SCHOOLING
J. exp. Biol. 175, 325 331 (1993) Printed in Great Britain The Company of Biologists Limited 1993 325 SHORT COMMUNICATION AN ANNULAR RESPIROMETER FOR MEASURING AEROBIC METABOLIC RATES OF LARGE, SCHOOLING
Within Phase I, researchers have identified four tasks that they think are essential for designing the most informative study.
 Revised Study Plan June 30, 2005 I. Introduction The Merwin upstream fish passage facility was originally completed with construction of the Merwin Dam. This trapping facility historically operated with
Revised Study Plan June 30, 2005 I. Introduction The Merwin upstream fish passage facility was originally completed with construction of the Merwin Dam. This trapping facility historically operated with
SALMON FACTS. Chinook Salmon. Oncorhynchus tshawytscha
 SALMON FACTS Chinook Salmon Oncorhynchus tshawytscha Other names: king, tyee, blackmouth (immature) Average size: 10-15 lbs, up to 135 lbs Fall spawner; fall, spring, and summer runs Chinook salmon are
SALMON FACTS Chinook Salmon Oncorhynchus tshawytscha Other names: king, tyee, blackmouth (immature) Average size: 10-15 lbs, up to 135 lbs Fall spawner; fall, spring, and summer runs Chinook salmon are
ACUTE TEMPERATURE TOLERANCE OF JUVENILE CHINOOK SALMON FROM THE MOKELUMNE RIVER
 ACUTE TEMPERATURE TOLERANCE OF JUVENILE CHINOOK SALMON FROM THE MOKELUMNE RIVER Charles H. Hanson, Ph.D. Hanson Environmental, Inc. SUMMARY A series of static acute tests were performed to determine the
ACUTE TEMPERATURE TOLERANCE OF JUVENILE CHINOOK SALMON FROM THE MOKELUMNE RIVER Charles H. Hanson, Ph.D. Hanson Environmental, Inc. SUMMARY A series of static acute tests were performed to determine the
Fish Passage Studies I: Sea Lamprey Behaviour During Negotaiton of Technical and Nature-Like Fish Passes
 University of Massachusetts - Amherst ScholarWorks@UMass Amherst International Conference on Engineering and Ecohydrology for Fish Passage International Conference on Engineering and Ecohydrology for Fish
University of Massachusetts - Amherst ScholarWorks@UMass Amherst International Conference on Engineering and Ecohydrology for Fish Passage International Conference on Engineering and Ecohydrology for Fish
RELEASES OF HIGH-SEAS SALMON TAGS BY U.S. VESSELS IN 2006
 NPAFC Doc. 1038 Rev. Rev. Date: RELEASES OF HIGH-SEAS SALMON TAGS BY U.S. VESSELS IN 2006 James M. Murphy 1, Robert V. Walker 2, Jamal H. Moss 1, Kristin Cieciel 1, Nancy D. Davis 2, Katherine W. Myers
NPAFC Doc. 1038 Rev. Rev. Date: RELEASES OF HIGH-SEAS SALMON TAGS BY U.S. VESSELS IN 2006 James M. Murphy 1, Robert V. Walker 2, Jamal H. Moss 1, Kristin Cieciel 1, Nancy D. Davis 2, Katherine W. Myers
Session B9: Size Matters Even for the Ubiquitous Fish Speed Metric of BL/S
 University of Massachusetts - Amherst ScholarWorks@UMass Amherst International Conference on Engineering and Ecohydrology for Fish Passage International Conference on Engineering and Ecohydrology for Fish
University of Massachusetts - Amherst ScholarWorks@UMass Amherst International Conference on Engineering and Ecohydrology for Fish Passage International Conference on Engineering and Ecohydrology for Fish
EFFECTS OF IMPORT AND INVENTORY AMOUNTS ON CHANGES IN WHOLESALE PRICES OF SALMON IN JAPAN
 EFFECTS OF IMPORT AND INVENTORY AMOUNTS ON CHANGES IN WHOLESALE PRICES OF SALMON IN JAPAN Ikutaro Shimizu National Salmon Resources Center, ikutaro@salmon.affrc.go.jp ABSTRACT Japanese salmon fishery is
EFFECTS OF IMPORT AND INVENTORY AMOUNTS ON CHANGES IN WHOLESALE PRICES OF SALMON IN JAPAN Ikutaro Shimizu National Salmon Resources Center, ikutaro@salmon.affrc.go.jp ABSTRACT Japanese salmon fishery is
Journal of Fish Biology (2009) 74, doi: /j x, available online at
 74, doi: /j x, available online at") Journal of Fish Biology (29) 74, 1323 1336 doi:1.1111/j.195-8649.29.2213.x, available online at http://www.blackwell-synergy.com Physiological, energetic and behavioural correlates of successful fishway
Journal of Fish Biology (29) 74, 1323 1336 doi:1.1111/j.195-8649.29.2213.x, available online at http://www.blackwell-synergy.com Physiological, energetic and behavioural correlates of successful fishway
Corrigendum. For GC sockeye salmon: scope for activity=11.22/{1+[(t 16.63)/8.11] 2 }.
![Corrigendum. For GC sockeye salmon: scope for activity=11.22/{1+[(t 16.63)/8.11] 2 }.](/thumbs/87/95742933.jpg "Corrigendum. For GC sockeye salmon: scope for activity=11.22/{1+[(t 16.63)/8.11] 2 }.") Corrigendum Lee, C. G., Farrell, A. P., Lotto, A., MacNutt, M. J., Hinch, S. G. and Healey, M. C. (23). The effect of temperature on swimming performance and oxygen consumption in adult sockeye (Oncorhynchus
Corrigendum Lee, C. G., Farrell, A. P., Lotto, A., MacNutt, M. J., Hinch, S. G. and Healey, M. C. (23). The effect of temperature on swimming performance and oxygen consumption in adult sockeye (Oncorhynchus
BOGUS CREEK SALMON STUDIES 2002
 BOGUS CREEK SALMON STUDIES 2002 BY: JEANNINE RICHEY California Department of Fish and Game KLAMATH RIVER PROJECT 303 SOUTH STREET YREKA, CALIFORNIA 96097 (530) 842-3109 California Department of Fish and
BOGUS CREEK SALMON STUDIES 2002 BY: JEANNINE RICHEY California Department of Fish and Game KLAMATH RIVER PROJECT 303 SOUTH STREET YREKA, CALIFORNIA 96097 (530) 842-3109 California Department of Fish and
Occurrence of Myxobolus arcticus and M. neurobius (Myxozoa: Myxosporea) in Masu Salmon Oncorhynchus masou from Northern Japan
 in Masu Salmon Oncorhynchus masou from Northern Japan") Scientific Reports of the Hokkaido Salmon Hatchery No. 49: 35-, March 1995 Occurrence of Myxobolus arcticus and M. neurobius (Myxozoa: Myxosporea) in Masu Salmon Oncorhynchus masou from Northern Japan
Scientific Reports of the Hokkaido Salmon Hatchery No. 49: 35-, March 1995 Occurrence of Myxobolus arcticus and M. neurobius (Myxozoa: Myxosporea) in Masu Salmon Oncorhynchus masou from Northern Japan
From: Michael A. Jepson, Tami C. Clabough, and Christopher C. Caudill
 To: Derek Fryer, USACE Walla Walla District From: Michael A. Jepson, Tami C. Clabough, and Christopher C. Caudill RE: Evaluation of Radio-tagged Adult Chinook Salmon Behavior in Response to Nighttime Flow
To: Derek Fryer, USACE Walla Walla District From: Michael A. Jepson, Tami C. Clabough, and Christopher C. Caudill RE: Evaluation of Radio-tagged Adult Chinook Salmon Behavior in Response to Nighttime Flow
THE EFFECTS OF REDUCED GILL AREA AND HYPEROXIA ON THE OXYGEN CONSUMPTION AND SWIMMING SPEED OF RAINBOW TROUT
 J. exp. Biol. 127, 349-354 (1987) 349 f Great Britain The Company of Biologists Limited 1987 THE EFFECTS OF REDUCED GILL AREA AND HYPEROXIA ON THE OXYGEN CONSUMPTION AND SWIMMING SPEED OF RAINBOW TROUT
J. exp. Biol. 127, 349-354 (1987) 349 f Great Britain The Company of Biologists Limited 1987 THE EFFECTS OF REDUCED GILL AREA AND HYPEROXIA ON THE OXYGEN CONSUMPTION AND SWIMMING SPEED OF RAINBOW TROUT
Salmon age and size at maturity: Patterns and processes
 Salmon age and size at maturity: Patterns and processes 1. Age Designation 2. Variation among populations 1. Latitude 2. Within regions 3. Within watersheds 3. Variation within populations 1. Smolt size
Salmon age and size at maturity: Patterns and processes 1. Age Designation 2. Variation among populations 1. Latitude 2. Within regions 3. Within watersheds 3. Variation within populations 1. Smolt size
The Life History and Management of Alewife (Alosa pseudoharengus)
") The Life History and Management of Alewife (Alosa pseudoharengus) Abigail Franklin, University of Massachusetts, Amherst Department of Natural Resources Conservation The Biology and Management of Alewife
The Life History and Management of Alewife (Alosa pseudoharengus) Abigail Franklin, University of Massachusetts, Amherst Department of Natural Resources Conservation The Biology and Management of Alewife
Ecology of Columbia River redband trout (Oncorhynchus mykiss gairdneri) in high desert streams
 in high desert streams") Ecology of Columbia River redband trout (Oncorhynchus mykiss gairdneri) in high desert streams Dry Creek October 2012 Chris A. Walser Ph.D., Professor of Biology Department of Biology and Environmental
Ecology of Columbia River redband trout (Oncorhynchus mykiss gairdneri) in high desert streams Dry Creek October 2012 Chris A. Walser Ph.D., Professor of Biology Department of Biology and Environmental
FITFISH 2016 Swimming Respirometry. Alina Antache Caroline Navjord Núria Vila İlkay Hüyüklü
 FITFISH 2016 Swimming Respirometry Alina Antache Caroline Navjord Núria Vila İlkay Hüyüklü Index Preparing for the experiments Sea Bass Experiments and Results Pompano Experiments and Results Conclusions
FITFISH 2016 Swimming Respirometry Alina Antache Caroline Navjord Núria Vila İlkay Hüyüklü Index Preparing for the experiments Sea Bass Experiments and Results Pompano Experiments and Results Conclusions
(breed) Anadromous. wholly seawater
 Anadromous. wholly seawater") Maintenance of Constancy of the Internal Environment during Fish Migration Between Fresh and Seawater Group 4: Kuan Yi Xian, Jeeann Lee Tien Ming Lee Tze Chuen Lee Wan-Jean Lee Yee Tjin Migration Seasonal
Maintenance of Constancy of the Internal Environment during Fish Migration Between Fresh and Seawater Group 4: Kuan Yi Xian, Jeeann Lee Tien Ming Lee Tze Chuen Lee Wan-Jean Lee Yee Tjin Migration Seasonal
Smolt Monitoring Protocol at COE Dams On the Lower Snake and Lower Columbia rivers
 Smolt Monitoring Protocol at COE Dams On the Lower Snake and Lower Columbia rivers 1.0 Introduction There are two primary goals of the Smolt Monitoring Program (SMP); to provide realtime data on juvenile
Smolt Monitoring Protocol at COE Dams On the Lower Snake and Lower Columbia rivers 1.0 Introduction There are two primary goals of the Smolt Monitoring Program (SMP); to provide realtime data on juvenile
Upstream Passage Assessment of American Shad Using 3D Acoustic Telemetry
 Upstream Passage Assessment of American Shad Using 3D Acoustic Telemetry Timothy Hogan, Alden Research Laboratory Corey Wright, Blue Leaf Environmental Skip Medford, Enel Green Power North America Abstract
Upstream Passage Assessment of American Shad Using 3D Acoustic Telemetry Timothy Hogan, Alden Research Laboratory Corey Wright, Blue Leaf Environmental Skip Medford, Enel Green Power North America Abstract
BULL TROUT OPERATIONAL PLAN
 BULL TROUT OPERATIONAL PLAN ROCKY REACH HYDROELECTRIC PROJECT FERC Project No. 2145 April 10, 2001 Public Utility District No. 1 of Chelan County Wenatchee, Washington BULL TROUT OPERATIONAL PLAN This
BULL TROUT OPERATIONAL PLAN ROCKY REACH HYDROELECTRIC PROJECT FERC Project No. 2145 April 10, 2001 Public Utility District No. 1 of Chelan County Wenatchee, Washington BULL TROUT OPERATIONAL PLAN This
ENTRANCES USED AND PASSAGE THROUGH FISHWAYS FOR ADULT CHINOOK SALMON AND STEELHEAD
 ENTRANCES USED AND PASSAGE THROUGH FISHWAYS FOR ADULT CHINOOK SALMON AND STEELHEAD Part III of Final Report for MIGRATION OF ADULT CHINOOK SALMON AND STEELHEAD PAST DAMS AND THROUGH RESERVOIRS IN THE LOWER
ENTRANCES USED AND PASSAGE THROUGH FISHWAYS FOR ADULT CHINOOK SALMON AND STEELHEAD Part III of Final Report for MIGRATION OF ADULT CHINOOK SALMON AND STEELHEAD PAST DAMS AND THROUGH RESERVOIRS IN THE LOWER
"Recommended Improvements for the Next Pacific Salmon Treaty"
 "Recommended Improvements for the Next Pacific Salmon Treaty" Randall M. Peterman School of Resource and Environmental Management Simon Fraser University Burnaby, British Columbia, Canada Web site: http://www.rem.sfu.ca/fishgrp/
"Recommended Improvements for the Next Pacific Salmon Treaty" Randall M. Peterman School of Resource and Environmental Management Simon Fraser University Burnaby, British Columbia, Canada Web site: http://www.rem.sfu.ca/fishgrp/
SCIENTIFIC COMMITTEE SEVENTH REGULAR SESSION August 2011 Pohnpei, Federated States of Micronesia
 SCIENTIFIC COMMITTEE SEVENTH REGULAR SESSION 9-17 August 2011 Pohnpei, Federated States of Micronesia CPUE of skipjack for the Japanese offshore pole and line using GPS and catch data WCPFC-SC7-2011/SA-WP-09
SCIENTIFIC COMMITTEE SEVENTH REGULAR SESSION 9-17 August 2011 Pohnpei, Federated States of Micronesia CPUE of skipjack for the Japanese offshore pole and line using GPS and catch data WCPFC-SC7-2011/SA-WP-09
UNIT 4E. SALMON SUPPLEMENTAL INFORMATION
 UNIT 4E. SALMON SUPPLEMENTAL INFORMATION In the STUDENT HANDOUTS/TEACHER PAGES Section of this unit, we have provided you with a host of information from various sources to supplement not only this unit,
UNIT 4E. SALMON SUPPLEMENTAL INFORMATION In the STUDENT HANDOUTS/TEACHER PAGES Section of this unit, we have provided you with a host of information from various sources to supplement not only this unit,
Long-distance downstream movements by homing adult chinook salmon
 Journal of Fish Biology (2006) 68, 944 950 doi:10.1111/j.1095-8649.2006.00958.x, available online at http://www.blackwell-synergy.com Long-distance downstream movements by homing adult chinook salmon M.
Journal of Fish Biology (2006) 68, 944 950 doi:10.1111/j.1095-8649.2006.00958.x, available online at http://www.blackwell-synergy.com Long-distance downstream movements by homing adult chinook salmon M.
Fish Tech Weekly Outline January 14-18
 Fish Tech Weekly Outline January 14-18 TOPICS: salmon, trout, and char in Southeast Alaska salmonid identification overview salmonid life cycle and stages salmonid freshwater and marine distribution/residence
Fish Tech Weekly Outline January 14-18 TOPICS: salmon, trout, and char in Southeast Alaska salmonid identification overview salmonid life cycle and stages salmonid freshwater and marine distribution/residence
Judd Lake Adult Sockeye Salmon Data Report 2012
 Judd Lake Adult Sockeye Salmon Data Report 2012 Prepared by: Nathan Weber, Biologist July 2013 The Judd Lake Project was made possible through a State of Alaska Designated Legislative Grant. This page
Judd Lake Adult Sockeye Salmon Data Report 2012 Prepared by: Nathan Weber, Biologist July 2013 The Judd Lake Project was made possible through a State of Alaska Designated Legislative Grant. This page
Case Studies in Ecology and Evolution
 3 Applied population dynamics: managing Pacific salmon By the end of this chapter you should be able to: Estimate population size from simple mark-recapture data Construct stock-recruit graphs that relate
3 Applied population dynamics: managing Pacific salmon By the end of this chapter you should be able to: Estimate population size from simple mark-recapture data Construct stock-recruit graphs that relate
Jason Blackburn, Paul Hvenegaard, Dave Jackson, Tyler Johns, Chad Judd, Scott Seward and Juanna Thompson
 Alberta Conservation Association (ACA) Date: 2014-2015 Project Name: Owl River Walleye and Aquatic Habitat Assessment Fisheries Program Manager: Peter Aku Project Leader: Tyler Johns Primary ACA staff
Alberta Conservation Association (ACA) Date: 2014-2015 Project Name: Owl River Walleye and Aquatic Habitat Assessment Fisheries Program Manager: Peter Aku Project Leader: Tyler Johns Primary ACA staff
Technical Report Draft
 Technical Report 2004-11 Draft An Evaluation of Adult Chinook Salmon and Steelhead Behavior at Counting Windows of Mcary Dam during 2002 & 2003 and the orth Shore Counting Window at Ice Harbor Dam during
Technical Report 2004-11 Draft An Evaluation of Adult Chinook Salmon and Steelhead Behavior at Counting Windows of Mcary Dam during 2002 & 2003 and the orth Shore Counting Window at Ice Harbor Dam during
A COMPARISON OF THERMAL POLYGONS FOR BRITISH FRESHWATER TELEOSTS
 178 A. ELLIOTT A COMPARISON OF THERMAL POLYGONS FOR BRITISH FRESHWATER TELEOSTS Introduction ALEX ELLIOTT (J. A. Elliott, 51312 Muirhead House, University of Stirling, Stirling FK9 4LG, Scotland.) Almost
178 A. ELLIOTT A COMPARISON OF THERMAL POLYGONS FOR BRITISH FRESHWATER TELEOSTS Introduction ALEX ELLIOTT (J. A. Elliott, 51312 Muirhead House, University of Stirling, Stirling FK9 4LG, Scotland.) Almost
January 2016 Recent studies have shown that, in British Columbia regardless of the presence or
 Recent studies have shown that, in British Columbia regardless of the presence or absence of salmon farms, there is wide variability in sea lice prevalence in coastal locations. Sea lice are naturally-occurring
Recent studies have shown that, in British Columbia regardless of the presence or absence of salmon farms, there is wide variability in sea lice prevalence in coastal locations. Sea lice are naturally-occurring
Executive Summary. Map 1. The Santa Clara River watershed with topography.
 Santa Clara River Steelhead Trout: Assessment and Recovery Opportunities December 2005 Prepared By: Matt Stoecker and Elise Kelley, Ph.D. University of California, Santa Barbara Prepared For: The Santa
Santa Clara River Steelhead Trout: Assessment and Recovery Opportunities December 2005 Prepared By: Matt Stoecker and Elise Kelley, Ph.D. University of California, Santa Barbara Prepared For: The Santa
Assessment of an International Breaststroke Swimmer Using a Race Readiness Test
 International Journal of Sports Physiology and Performance, 2009, 4, 139-143 2009 Human Kinetics, Inc. Assessment of an International Breaststroke Swimmer Using a Race Readiness Test Kevin G. Thompson
International Journal of Sports Physiology and Performance, 2009, 4, 139-143 2009 Human Kinetics, Inc. Assessment of an International Breaststroke Swimmer Using a Race Readiness Test Kevin G. Thompson
Emily M. Standen, Scott G. Hinch, Michael C. Healey, and Anthony P. Farrell
 1809 Energetic costs of migration through the Fraser River Canyon, British Columbia, in adult pink (Oncorhynchus gorbuscha) and sockeye (Oncorhynchus nerka) salmon as assessed by EMG telemetry Emily M.
1809 Energetic costs of migration through the Fraser River Canyon, British Columbia, in adult pink (Oncorhynchus gorbuscha) and sockeye (Oncorhynchus nerka) salmon as assessed by EMG telemetry Emily M.
Monitoring of Downstream Fish Passage at Cougar Dam in the South Fork McKenzie River, Oregon February 8, By Greg A.
 Monitoring of Downstream Fish Passage at Cougar Dam in the South Fork McKenzie River, Oregon 1998- February 8, 2 By Greg A. Taylor Oregon Dept. of Fish & Wildlife 315 E. Main Street Springfield, OR 97478
Monitoring of Downstream Fish Passage at Cougar Dam in the South Fork McKenzie River, Oregon 1998- February 8, 2 By Greg A. Taylor Oregon Dept. of Fish & Wildlife 315 E. Main Street Springfield, OR 97478
Early Marine Migrations. General geography Four general patterns Influence of genetics
 Early Marine Migrations General geography Four general patterns Influence of genetics Coastal range of anadromous Pacific salmon and trout Techniques employed to study salmon at sea Recently developed
Early Marine Migrations General geography Four general patterns Influence of genetics Coastal range of anadromous Pacific salmon and trout Techniques employed to study salmon at sea Recently developed
Technical Report 99-6 EVALUATION OF RUNNING TURBINE 1 AT MAXIMUM CAPACITY ON PASSAGE OF ADULT SALMON AND STEELHEAD AT JOHN DAY DAM
 Technical Report 99-6 EVALUATION OF RUNNING TURBINE 1 AT MAXIMUM CAPACITY ON PASSAGE OF ADULT SALMON AND STEELHEAD AT JOHN DAY DAM - 1997 A report for Project MPE-P-95-1 by T.C. Bjornn, C.A. Peery, K.R.
Technical Report 99-6 EVALUATION OF RUNNING TURBINE 1 AT MAXIMUM CAPACITY ON PASSAGE OF ADULT SALMON AND STEELHEAD AT JOHN DAY DAM - 1997 A report for Project MPE-P-95-1 by T.C. Bjornn, C.A. Peery, K.R.
Striped Bass and White Hybrid (x) Striped Bass Management and Fishing in Pennsylvania
 Striped Bass Management and Fishing in Pennsylvania") Striped Bass and White Hybrid (x) Striped Bass Management and Fishing in Pennsylvania Prepared by R. Lorantas, D. Kristine and C. Hobbs PFBC Warmwater Unit 2005 (stocking numbers updated after 2005) Goal:
Striped Bass and White Hybrid (x) Striped Bass Management and Fishing in Pennsylvania Prepared by R. Lorantas, D. Kristine and C. Hobbs PFBC Warmwater Unit 2005 (stocking numbers updated after 2005) Goal:
2013 Electrofishing Program Summary. Miramichi Salmon Association In collaboration with the Department of Fisheries and Oceans
 2013 Electrofishing Program Summary Miramichi Salmon Association In collaboration with the Department of Fisheries and Oceans Prepared by: Alex Parker Biologist Miramichi Salmon Association 485, Route
2013 Electrofishing Program Summary Miramichi Salmon Association In collaboration with the Department of Fisheries and Oceans Prepared by: Alex Parker Biologist Miramichi Salmon Association 485, Route
Technical Report
 Technical Report 2003-04 Effects of the Shad Fishery on Passage of Adult Chinook Salmon through the Oregon-shore Fishway Ladder at The Dalles Dam 2002 M. A. Jepson, C.L. Williams, and C.A. Peery Idaho
Technical Report 2003-04 Effects of the Shad Fishery on Passage of Adult Chinook Salmon through the Oregon-shore Fishway Ladder at The Dalles Dam 2002 M. A. Jepson, C.L. Williams, and C.A. Peery Idaho
Meaghan J. MacNutt, Scott G. Hinch, Chris G. Lee, James R. Phibbs, Andrew G. Lotto, Michael C. Healey, and Anthony P. Farrell
 88 Temperature effects on swimming performance, energetics, and aerobic capacities of mature adult pink salmon (Oncorhynchus gorbuscha) compared with those of sockeye salmon (Oncorhynchus nerka) Meaghan
88 Temperature effects on swimming performance, energetics, and aerobic capacities of mature adult pink salmon (Oncorhynchus gorbuscha) compared with those of sockeye salmon (Oncorhynchus nerka) Meaghan
Spilling Water at Hydroelectric Projects in the Columbia and Snake Rivers How Does It Benefit Salmon?
 Spilling Water at Hydroelectric Projects in the Columbia and Snake Rivers How Does It Benefit Salmon? Hydropower development in the Columbia and Snake rivers has left its mark on salmonid populations,
Spilling Water at Hydroelectric Projects in the Columbia and Snake Rivers How Does It Benefit Salmon? Hydropower development in the Columbia and Snake rivers has left its mark on salmonid populations,
Patterns of migration and delay observed in Summer Steelhead from the Upper Columbia and Snake River Basins from PIT tag data
 West Coast Region Patterns of migration and delay observed in Summer Steelhead from the Upper Columbia and Snake River Basins from PIT tag data Blane Bellerud August, 2015 Columbia River Basin Columbia
West Coast Region Patterns of migration and delay observed in Summer Steelhead from the Upper Columbia and Snake River Basins from PIT tag data Blane Bellerud August, 2015 Columbia River Basin Columbia
Chinook salmon (photo by Roger Tabor)
") Stream Residence 1. The choice : emigration vs. residence 2. Food: Abundance, density, and variation 3. Territory and Dominance 4. Fish Responses: Distribution, growth, survival 5. Mortality Migration
Stream Residence 1. The choice : emigration vs. residence 2. Food: Abundance, density, and variation 3. Territory and Dominance 4. Fish Responses: Distribution, growth, survival 5. Mortality Migration
Homing and Straying, Part II Mechanisms of Homing: Imprinting, Genetics, and Pheromones
 Homing and Straying, Part II Mechanisms of Homing: Imprinting, Genetics, and Pheromones Deepwater Bay sockeye experiment Olfactory nerve cut Tended to stay at release site Moved toward Fraser R. but did
Homing and Straying, Part II Mechanisms of Homing: Imprinting, Genetics, and Pheromones Deepwater Bay sockeye experiment Olfactory nerve cut Tended to stay at release site Moved toward Fraser R. but did
A REVIEW AND EVALUATION OF NATURAL MORTALITY FOR THE ASSESSMENT AND MANAGEMENT OF YELLOWFIN TUNA IN THE EASTERN PACIFIC OCEAN
 A REVIEW AND EVALUATION OF NATURAL MORTALITY FOR THE ASSESSMENT AND MANAGEMENT OF YELLOWFIN TUNA IN THE EASTERN PACIFIC OCEAN Mark N. Maunder and Alex Aires-da-Silva Outline YFT history Methods to estimate
A REVIEW AND EVALUATION OF NATURAL MORTALITY FOR THE ASSESSMENT AND MANAGEMENT OF YELLOWFIN TUNA IN THE EASTERN PACIFIC OCEAN Mark N. Maunder and Alex Aires-da-Silva Outline YFT history Methods to estimate
The Role of the NPAFC in Conservation and Protection of Pacific Salmon
 The Role of the NPAFC in Conservation and Protection of Pacific Salmon Vladimir Fedorenko Executive Director and Shigehiko Urawa Deputy Director North Pacific Anadromous Fish Commission Vancouver, Canada
The Role of the NPAFC in Conservation and Protection of Pacific Salmon Vladimir Fedorenko Executive Director and Shigehiko Urawa Deputy Director North Pacific Anadromous Fish Commission Vancouver, Canada
Five Counties Salmonid Conservation Program - Fish Passage Design Workshop. February 2013
 Program - Aquatic Organisms and Stream Crossings Ecological Connectivity A watershed is a network of channels that drain a common boundary. Channel characteristics formed by interaction of precipitation,
Program - Aquatic Organisms and Stream Crossings Ecological Connectivity A watershed is a network of channels that drain a common boundary. Channel characteristics formed by interaction of precipitation,
Susitna-Watana Hydroelectric Project (FERC No ) Salmon Escapement Study Study Plan Section 9.7
 Salmon Escapement Study Study Plan Section 9.7") (FERC No. 14241) Salmon Escapement Study Study Plan Section 9.7 Part D: Supplemental Information to June 2014 Initial Study Report Prepared for Prepared by LGL Alaska Research Associates, Inc. & Alaska
(FERC No. 14241) Salmon Escapement Study Study Plan Section 9.7 Part D: Supplemental Information to June 2014 Initial Study Report Prepared for Prepared by LGL Alaska Research Associates, Inc. & Alaska
MINISTRY OF ENVIRONMENT ENVIRONMENTAL STEWARDSHIP DIVISION FISH AND WILDLIFE BRANCH. Horsefly River Angling Management Plan
 MINISTRY OF ENVIRONMENT ENVIRONMENTAL STEWARDSHIP DIVISION FISH AND WILDLIFE BRANCH Horsefly River Angling Management Plan January 30 th, 2006 Area/Fishery Description The Horsefly River watershed provides
MINISTRY OF ENVIRONMENT ENVIRONMENTAL STEWARDSHIP DIVISION FISH AND WILDLIFE BRANCH Horsefly River Angling Management Plan January 30 th, 2006 Area/Fishery Description The Horsefly River watershed provides
Abundance of Steelhead and Coho Salmon in the Lagunitas Creek Drainage, Marin County, California
 scanned for KRIS Abundance of Steelhead and Coho Salmon in the Lagunitas Creek Drainage, Marin County, California Prepared for: Marin Municipal Water District 220 Nellen Drive Corte Madera, California
scanned for KRIS Abundance of Steelhead and Coho Salmon in the Lagunitas Creek Drainage, Marin County, California Prepared for: Marin Municipal Water District 220 Nellen Drive Corte Madera, California
A THESIS M.S. CIVIL ENGINEERING UNIVERSITY OF ALASKA ANCHORAGE ALEXANDRA WEST JEFFERIES, P.E.
 Assessing Fish Passage Success in Culvert Structures with the Development of a Two-Dimensional Algorithm Considering Physical Capabilities of Juvenile Salmonids ALEXANDRA WEST JEFFERIES, P.E. A THESIS
Assessing Fish Passage Success in Culvert Structures with the Development of a Two-Dimensional Algorithm Considering Physical Capabilities of Juvenile Salmonids ALEXANDRA WEST JEFFERIES, P.E. A THESIS
SEA GRANT PROGRESS REPORT
 SEA GRANT PROGRESS REPORT Project Title: The relationship between seasonal migrations of berried female lobster Homarus americanus, egg development and larval survival. Principal Investigator: Winsor H.
SEA GRANT PROGRESS REPORT Project Title: The relationship between seasonal migrations of berried female lobster Homarus americanus, egg development and larval survival. Principal Investigator: Winsor H.
Okanagan Sockeye Reintroduction
 Okanagan Sockeye Reintroduction Backgrounder Elders accounts, and other forms of Traditional Ecological Knowledge (TEK) tell us that many species of salmon once came into the Okanagan Valley and tell us
Okanagan Sockeye Reintroduction Backgrounder Elders accounts, and other forms of Traditional Ecological Knowledge (TEK) tell us that many species of salmon once came into the Okanagan Valley and tell us
ADULT SALMON AND STEELHEAD PASSAGE THROUGH FISHWAYS AND TRANSITION POOLS AT JOHN DAY DAM, Report for project MPE-P-95-1
 Technical Report 2008-4 ADULT SALMON AND STEELHEAD PASSAGE THROUGH FISHWAYS AND TRANSITION POOLS AT JOHN DAY DAM, 1997-2001 Report for project MPE-P-95-1 M.L. Keefer, C.A. Peery, T.C. Bjornn, M.A. Jepson,
Technical Report 2008-4 ADULT SALMON AND STEELHEAD PASSAGE THROUGH FISHWAYS AND TRANSITION POOLS AT JOHN DAY DAM, 1997-2001 Report for project MPE-P-95-1 M.L. Keefer, C.A. Peery, T.C. Bjornn, M.A. Jepson,
EXPERIMENTAL STUDY ON SNOW BEHAVIOR AROUND FENCES INSTALLED ALONG ELEVATED HIGHWAY
 ISTP-16, 25, PRAGUE 16 TH INTERNATIONAL SYMPOSIUM ON TRANSPORT PHENOMENA EXPERIMENTAL STUDY ON SNOW BEHAVIOR AROUND FENCES INSTALLED ALONG ELEVATED HIGHWAY Akinori Nakata*, Haruo Soeda*, Junji Onishi*,
ISTP-16, 25, PRAGUE 16 TH INTERNATIONAL SYMPOSIUM ON TRANSPORT PHENOMENA EXPERIMENTAL STUDY ON SNOW BEHAVIOR AROUND FENCES INSTALLED ALONG ELEVATED HIGHWAY Akinori Nakata*, Haruo Soeda*, Junji Onishi*,
Red Salmon Lake Data Report 2011
 Red Salmon Lake Data Report 2011 Prepared by: Nathan Weber, Biologist February 2012 The Red Salmon Lake Project was made possible through an Alaskan Sustainable Salmon Fund grant received from the Alaska
Red Salmon Lake Data Report 2011 Prepared by: Nathan Weber, Biologist February 2012 The Red Salmon Lake Project was made possible through an Alaskan Sustainable Salmon Fund grant received from the Alaska
Proposed 2018 Fisheries Management Measures to Support Recovery of Interior Fraser River Steelhead
 Proposed 2018 Fisheries Management Measures to Support Recovery of Interior Fraser River Steelhead 22-March-2018 Spawning escapements of two Interior Fraser River steelhead stocks, Thompson and Chilcotin
Proposed 2018 Fisheries Management Measures to Support Recovery of Interior Fraser River Steelhead 22-March-2018 Spawning escapements of two Interior Fraser River steelhead stocks, Thompson and Chilcotin
2010 Lower Fraser River Sockeye Recreational Hook and Release Mortality Study
 2010 Lower Fraser River Sockeye Recreational Hook and Release Mortality Study Summary of Findings Prepared by: J. O. Thomas & Associates Ltd. Prepared for: Debra Sneddon, FOC Fraser River Area 18-November-2010
2010 Lower Fraser River Sockeye Recreational Hook and Release Mortality Study Summary of Findings Prepared by: J. O. Thomas & Associates Ltd. Prepared for: Debra Sneddon, FOC Fraser River Area 18-November-2010
Study Update Fish Distribution and Species Composition
 Study Update Fish Distribution and Species Composition Goals and Study Area Gain a better understanding of fish species composition and abundance in waters associated with the Packwood Lake Hydroelectric
Study Update Fish Distribution and Species Composition Goals and Study Area Gain a better understanding of fish species composition and abundance in waters associated with the Packwood Lake Hydroelectric
RELEASES AND RECOVERIES OF U.S. SALMONID DATA STORAGE TAGS, AND RECOVERIES OF HIGH SEAS TAGS IN NORTH AMERICA, 2001
 NPAFC Doc. 556 Rev. RELEASES AND RECOVERIES OF U.S. SALMONID DATA STORAGE TAGS, AND RECOVERIES OF HIGH SEAS TAGS IN NORTH AMERICA, 2001 Robert V. Walker, Nancy D. Davis, and Katherine W. Myers University
NPAFC Doc. 556 Rev. RELEASES AND RECOVERIES OF U.S. SALMONID DATA STORAGE TAGS, AND RECOVERIES OF HIGH SEAS TAGS IN NORTH AMERICA, 2001 Robert V. Walker, Nancy D. Davis, and Katherine W. Myers University
R0204-NPAFC Salmon Tagging Progress Report: July 1, 2005
 Project #: R4 Title: NPAFC Salmon Tagging Principal Investigator and Recipient Organization: Jack Helle (jack.helle@noaa.gov), Chairman, BASIS Working Group; North Pacific Anadromous Fish Commission (NPAFC),
Project #: R4 Title: NPAFC Salmon Tagging Principal Investigator and Recipient Organization: Jack Helle (jack.helle@noaa.gov), Chairman, BASIS Working Group; North Pacific Anadromous Fish Commission (NPAFC),
ALASKA DEPARTMENT OF FISH AND GAME DIVISION OF COMMERCIAL FISHERIES NEWS RELEASE
 ALASKA DEPARTMENT OF FISH AND GAME DIVISION OF COMMERCIAL FISHERIES NEWS RELEASE Sam Cotten, Commissioner Scott Kelley, Director Contact: Cordova ADF&G Stormy Haught, PWS Finfish Research Biologist 401
ALASKA DEPARTMENT OF FISH AND GAME DIVISION OF COMMERCIAL FISHERIES NEWS RELEASE Sam Cotten, Commissioner Scott Kelley, Director Contact: Cordova ADF&G Stormy Haught, PWS Finfish Research Biologist 401
Attachment 2. Exhibit (I) Public Correspondence received as of May 25, 2018.
 Public Correspondence received as of May 25, 2018.") Attachment 2 Exhibit (I) Public Correspondence received as of May 25, 2018. From: To: Subject: Date: Attachments: Michelle Tate April H Mack FW: Thermal Refuge comments and recommendations Tuesday, March
Attachment 2 Exhibit (I) Public Correspondence received as of May 25, 2018. From: To: Subject: Date: Attachments: Michelle Tate April H Mack FW: Thermal Refuge comments and recommendations Tuesday, March
STUDY PERFORMANCE REPORT
 STUDY PERFORMANCE REPORT State: Michigan Project No.: F-80-R-7 Study No.: 230654 Title: Evaluation of brown trout and steelhead competitive interactions in Hunt Creek, Michigan. Period Covered: October
STUDY PERFORMANCE REPORT State: Michigan Project No.: F-80-R-7 Study No.: 230654 Title: Evaluation of brown trout and steelhead competitive interactions in Hunt Creek, Michigan. Period Covered: October
Date: 25 September Introduction
 To: David Clugston, USACE Portland District From: Matt Keefer, Eric Johnson, Tami Clabough, Mike Jepson, Chris Caudill, Mary Moser RE: Preliminary evaluation of radiotelemetry and half-duplex PIT tag data
To: David Clugston, USACE Portland District From: Matt Keefer, Eric Johnson, Tami Clabough, Mike Jepson, Chris Caudill, Mary Moser RE: Preliminary evaluation of radiotelemetry and half-duplex PIT tag data
Stock Assessment of Anadromous Salmonids, 2003 Report Number: OPSW-ODFW
 THE OREGON PLAN for Salmon and Watersheds Stock Assessment of Anadromous Salmonids, 3 Report Number: OPSW-ODFW-- The Oregon Department of Fish and Wildlife prohibits discrimination in all of its programs
THE OREGON PLAN for Salmon and Watersheds Stock Assessment of Anadromous Salmonids, 3 Report Number: OPSW-ODFW-- The Oregon Department of Fish and Wildlife prohibits discrimination in all of its programs
ELECTRO-FISHING REPORT 2016 UPPER TWEED
 ELECTRO-FISHING REPORT 2016 UPPER TWEED The electro-fishing programme carried out each summer by The Tweed Foundation is part of our management plan, which details the information that is required to manage
ELECTRO-FISHING REPORT 2016 UPPER TWEED The electro-fishing programme carried out each summer by The Tweed Foundation is part of our management plan, which details the information that is required to manage
Alberta Conservation Association 2009/10 Project Summary Report. Project Name: North Saskatchewan and Ram Rivers Bull Trout Spawning Stock Assessment
 Alberta Conservation Association 2009/10 Project Summary Report Project Name: North Saskatchewan and Ram Rivers Bull Trout Spawning Stock Assessment Fisheries Program Manager: Peter Aku Project Leader:
Alberta Conservation Association 2009/10 Project Summary Report Project Name: North Saskatchewan and Ram Rivers Bull Trout Spawning Stock Assessment Fisheries Program Manager: Peter Aku Project Leader:
Sergey Zolotukhin, Ph.D., Khabarovsk TINRO Pacific Salmon Research Laboratory
 Sergey Zolotukhin, Ph.D., Khabarovsk TINRO Pacific Salmon Research Laboratory Trends in Amur River fall chum abundance and climate change from 1907-2008, faced to fishery management Transboundary river
Sergey Zolotukhin, Ph.D., Khabarovsk TINRO Pacific Salmon Research Laboratory Trends in Amur River fall chum abundance and climate change from 1907-2008, faced to fishery management Transboundary river
EVALUATION OF FISHWAY MODIFICATIONS TO IMPROVE PASSAGE OF ADULT CHINOOK SALMON AND STEELHEAD THROUGH THE TRANSITION POOL AT LOWER GRANITE DAM, 2008
 Technical Report 2009-3 EVALUATION OF FISHWAY MODIFICATIONS TO IMPROVE PASSAGE OF ADULT CHINOOK SALMON AND STEELHEAD THROUGH THE TRANSITION POOL AT LOWER GRANITE DAM, 2008 by T.S. Clabough, G.P. Naughton,
Technical Report 2009-3 EVALUATION OF FISHWAY MODIFICATIONS TO IMPROVE PASSAGE OF ADULT CHINOOK SALMON AND STEELHEAD THROUGH THE TRANSITION POOL AT LOWER GRANITE DAM, 2008 by T.S. Clabough, G.P. Naughton,
IDAHO COOPERATIVE FISH AND WILDLIFE RESEARCH UNIT
 Technical Report 211-2 IDAHO COOPERATIVE FISH AND WILDLIFE RESEARCH UNIT BEHAVIOR OF RADIO-TAGGED ADULT SPRING-SUMMER CHINOOK SALMON AT THE DALLES DAM IN RELATION TO SPILL VOLUME AND THE PRESENCE OF THE
Technical Report 211-2 IDAHO COOPERATIVE FISH AND WILDLIFE RESEARCH UNIT BEHAVIOR OF RADIO-TAGGED ADULT SPRING-SUMMER CHINOOK SALMON AT THE DALLES DAM IN RELATION TO SPILL VOLUME AND THE PRESENCE OF THE
Technical Report
 Technical Report 2004-2 An Evaluation of Adult Chinook Salmon and Steelhead Behavior at Counting Windows and through Vertical-slot Weirs of Bonneville Dam Using Radiotelemetry: 2001-2002 A Report for Study
Technical Report 2004-2 An Evaluation of Adult Chinook Salmon and Steelhead Behavior at Counting Windows and through Vertical-slot Weirs of Bonneville Dam Using Radiotelemetry: 2001-2002 A Report for Study
FISHERIES BLUE MOUNTAINS ADAPTATION PARTNERSHIP
 FISHERIES A warming climate, by itself, substantially affects the hydrology of watersheds in the Blue Mountains. Among the key hydrologic changes projected under all scenarios for the 2040s and beyond
FISHERIES A warming climate, by itself, substantially affects the hydrology of watersheds in the Blue Mountains. Among the key hydrologic changes projected under all scenarios for the 2040s and beyond
Appendix A Recommended EPA Temperature Thresholds for use in Establishing Thermal Potential and Species Life Stage Numeric Criteria
 Appendix A Recommended EPA Temperature Thresholds for use in Establishing Thermal Potential and Species Life Stage Numeric Criteria 1. Temperature Limits Recommended to Protect Salmonid Guilds In this
Appendix A Recommended EPA Temperature Thresholds for use in Establishing Thermal Potential and Species Life Stage Numeric Criteria 1. Temperature Limits Recommended to Protect Salmonid Guilds In this
Experiment of a new style oscillating water column device of wave energy converter
 http://www.aimspress.com/ AIMS Energy, 3(3): 421-427. DOI: 10.3934/energy.2015.3.421 Received date 16 April 2015, Accepted date 01 September 2015, Published date 08 September 2015 Research article Experiment
http://www.aimspress.com/ AIMS Energy, 3(3): 421-427. DOI: 10.3934/energy.2015.3.421 Received date 16 April 2015, Accepted date 01 September 2015, Published date 08 September 2015 Research article Experiment
Oregon Hatchery Research Center January 2014 David L. G. Noakes, Professor & Director
 Oregon Hatchery Research Center January 2014 David L. G. Noakes, Professor & Director Research Proposal Homing Homing behavior is a striking feature of the biology of Pacific salmon, and is the basis for
Oregon Hatchery Research Center January 2014 David L. G. Noakes, Professor & Director Research Proposal Homing Homing behavior is a striking feature of the biology of Pacific salmon, and is the basis for
What is causing declines in the annual returns of Fraser River sockeye salmon?
 What is causing declines in the annual returns of Fraser River sockeye salmon? Mike Lapointe, Catherine Michielsens; Pacific Salmon Commission Secretariat Sue Grant, Bronwyn MacDonald; Fisheries and Oceans
What is causing declines in the annual returns of Fraser River sockeye salmon? Mike Lapointe, Catherine Michielsens; Pacific Salmon Commission Secretariat Sue Grant, Bronwyn MacDonald; Fisheries and Oceans
***Please Note*** April 3, Dear advisory committee members:
 April 3, 29 Dear advisory committee members: The fifth meeting of the CHF advisory committee will be held April 13 in Grants Pass from 6:-8:3 PM, and the purpose of this document is to help committee members
April 3, 29 Dear advisory committee members: The fifth meeting of the CHF advisory committee will be held April 13 in Grants Pass from 6:-8:3 PM, and the purpose of this document is to help committee members
Juvenile Fish Travel Time and Survival a common currency for evaluating fish passage operations
 FISH PASSAGE CENTER 1827 NE 44 th Ave., Suite 240, Portland, OR 97213 Phone: (503) 230-4099 Fax: (503) 230-7559 http://www.fpc.org/ e-mail us at fpcstaff@fpc.org MEMORANDUM TO: Ed Bowles, ODFW FROM: Michele
FISH PASSAGE CENTER 1827 NE 44 th Ave., Suite 240, Portland, OR 97213 Phone: (503) 230-4099 Fax: (503) 230-7559 http://www.fpc.org/ e-mail us at fpcstaff@fpc.org MEMORANDUM TO: Ed Bowles, ODFW FROM: Michele
Ad Hoc Review Group IP(06)12 FINAL. Implementation Plan. European Union (Denmark)
12 FINAL. Implementation Plan. European Union (Denmark)") Ad Hoc Review Group IP(06)12 FINAL Implementation Plan European Union (Denmark) 1. Introduction 1.1 The National management Plan for Salmon in Denmark In 2004 the Danish Forest and Nature Agency in co-operation
Ad Hoc Review Group IP(06)12 FINAL Implementation Plan European Union (Denmark) 1. Introduction 1.1 The National management Plan for Salmon in Denmark In 2004 the Danish Forest and Nature Agency in co-operation
Population Structure
 Population Structure Elements of Population Evolution Reproductive isolation (homing to natal sites) Differential natural and sexual selection Heritable traits Results Genetically determined population
Population Structure Elements of Population Evolution Reproductive isolation (homing to natal sites) Differential natural and sexual selection Heritable traits Results Genetically determined population
IDAHO COOPERATIVE FISH AND WILDLIFE RESEARCH UNIT
 Technical Report 2007-1 IDAHO COOPERATIVE FISH AND WILDLIFE RESEARCH UNIT EVALUATION OF FISHWAY MODIFICATIONS TO IMPROVE PASSAGE OF ADULT CHINOOK SALMON AND STEELHEAD THROUGH THE TRANSITION POOL AT LOWER
Technical Report 2007-1 IDAHO COOPERATIVE FISH AND WILDLIFE RESEARCH UNIT EVALUATION OF FISHWAY MODIFICATIONS TO IMPROVE PASSAGE OF ADULT CHINOOK SALMON AND STEELHEAD THROUGH THE TRANSITION POOL AT LOWER
Masahide Kaeriyama. Introduction. Ocean Distribution and Biomass of Pacific Salmon. diverse pattern of life history, and chum salmon (0.
 Scientific Reports of the Hokkaido Salmon Hatchery No SO: 101-111, November 1996 Effects of Population Density and Habitat Environment on Life History Strategy and Migration of Juvenile Sockeye (Oncorhynchus
Scientific Reports of the Hokkaido Salmon Hatchery No SO: 101-111, November 1996 Effects of Population Density and Habitat Environment on Life History Strategy and Migration of Juvenile Sockeye (Oncorhynchus
Job 1. Title: Estimate abundance of juvenile trout and salmon.
 STUDY PERFORMANCE REPORT State: Michigan Project No.: F-53-R-13 Study No.: 461 Title: Population dynamics of juvenile rainbow trout and coho salmon in Lake Superior tributaries Period Covered: April 1,
STUDY PERFORMANCE REPORT State: Michigan Project No.: F-53-R-13 Study No.: 461 Title: Population dynamics of juvenile rainbow trout and coho salmon in Lake Superior tributaries Period Covered: April 1,
P/FR/SK/54 DE LEEUW, A. D. MAMIN RIVER STEELMEAD: A STUDY ON A LIMITED TAGGING CPOX c. 1 mm SMITHERS MAMIN RIVER STEELHEAD: A STUDY ON A LIMITED
 P/FR/SK/54 DE LEEUW, A. D. MAMIN RIVER STEELMEAD: A STUDY ON A LIMITED TAGGING CPOX c. 1 mm SMITHERS MAMIN RIVER STEELHEAD: A STUDY ON A LIMITED TAGGING STUDY UNDERTAKEN DURING WINTER, 1984 by A.D. de
P/FR/SK/54 DE LEEUW, A. D. MAMIN RIVER STEELMEAD: A STUDY ON A LIMITED TAGGING CPOX c. 1 mm SMITHERS MAMIN RIVER STEELHEAD: A STUDY ON A LIMITED TAGGING STUDY UNDERTAKEN DURING WINTER, 1984 by A.D. de