AN ABSTRACT OF THE THESIS OF
|
|
|
- Nathan Hudson
- 5 years ago
- Views:
Transcription
1 AN ABSTRACT OF THE THESIS OF Selene Fregosi for the degree of Master of Science in Wildlife Science presented on May 1, Title: Development and Evaluation of an Animal-borne Active Acoustic Tag to Conduct Minimally Invasive Behavioral Response Studies on Marine Mammals. Abstract approved: Holger Klinck Development of new research tools is needed to better understand the potential effects of a noisier ocean on individual and populations of marine mammals. Current behavioral response studies utilize ship-mounted sound sources to induce short-term noise-related behavioral responses in tagged animals. Combining the tag with the sound source into an animal-mounted active acoustic and motion sensing tag could potentially allow for long-term, more controlled and cost-effective behavioral response studies. Field tests were conducted on juvenile northern elephant seals, Mirounga angustirostris, using both natural and anthropogenic stimuli, to evaluate the potential of such a prototype tag. Results showed that such an instrument does elicit behavioral responses in tagged individuals. Responses during the ascending phase of a dive consisted of a dive inversion, with the animal diving as deep as or deeper than its original dive depth (seven of nine exposures). Change in dive depth following exposure was significantly larger than change in depth for non-exposure inversions. A single exposure at the bottom phase of a deep dive followed the same pattern. Dive inversions were observed following white noise, sperm whale clicks, killer whale whistles, and sonar exposures, but not following common dolphin whistles. Responses to exposures received during the descending phase of a dive resulted in an increased descent rate in nine of ten exposures. All eight exposures during shallow dives, where the animals were likely limited by bathymetry from diving any deeper, were characterize by increased flow noise following exposure, an indicator of increased swim speed. Results
2 showing differential responses to specific exposure stimuli were inconclusive. Tag improvements and additional field efforts are needed to validate the tag s use in behavioral response studies to specific acoustic stimuli. There is potential to use this technology to study physiological effects of extended deep dives on marine mammals as well as frequency dependent hearing because of its ability to induce prolonged, unplanned dives in response to man-made sounds.
3 Copyright by Selene Fregosi May 1, 2015 All Rights Reserved
4 Development and Evaluation of an Animal-borne Active Acoustic Tag to Conduct Minimally Invasive Behavioral Response Studies on Marine Mammals by Selene Fregosi A THESIS submitted to Oregon State University in partial fulfillment of the requirements for the degree of Master of Science Presented May 1, 2015 Commencement June 2015
5 Master of Science thesis of Selene Fregosi presented on May 1, 2015 APPROVED: Major Professor, representing Wildlife Science Head of the Department of Fisheries and Wildlife Dean of the Graduate School I understand that my thesis will become part of the permanent collection of Oregon State University libraries. My signature below authorizes release of my thesis to any reader upon request. Selene Fregosi, Author
6 ACKNOWLEDGEMENTS Many thanks to the faculty and staff of the Hatfield Marine Science Center (HMSC), the OSU Department of Fisheries and Wildlife, and NOAA s Pacific Marine Environmental Laboratory, for their guidance and support through this process. This research would not have been possible without the financial support from NOAA Fisheries, Office of Science and Technology, Ocean Acoustics Program (Jason Gedamke) and the Cooperative Institute for Marine Resources Studies (OSU/NOAA), and the assistance of several student volunteers at UC Santa Cruz. Completion of this thesis was additionally supported by the Markham First Year Award, Laurel s Block Grant Tuition Scholarship, OSU Provost s Graduate Fellowship, Mastin Travel Award, and Hatfield Student Organization Travel award. Thank you to my advisor, Holger Klinck, for thinking it was a good idea to take me on as a student. Thanks for letting me work on my dream project and for giving me the opportunity to become a Beaver! His trust in me to try new things, even when I didn t feel ready, pushed me to be the scientist I am today. Thank you to Markus Horning for supporting my student status, before all the logistics were worked out, for the expertise in pinniped diving behavior, and for the skiing. Thank you Brandon Southall, for the endless support since before OSU, for treating me like family while teaching me all there is to know about noise, marine mammals, and fileting fish. I owe special thanks to Sharon Nieukirk. You not only provided countless reads of ALL of my fellowship applications, posters, and this thesis, but also emergency chocolate, encouraging confidence boosts, and general life advice from someone I truly admire. To my friends and labmates at OSU and HMSC, especially Lizz Mulligan, Sophie Pierszalowski, Niki Diogou, Shea Steingass, and Michelle Fournet, I m so grateful you guys had done this before to be able to answer all my silly questions and help me make friends. LaTreese Denson, thanks for being my writing buddy; sharing library rooms and laugh breaks along the way. I could not have done any of this without my Santa Cruz family Colleen and Caro, you are my personal cheerleaders, my Seal
7 Force Three, and no one gives hugs like you two. And Montana friends/family, thanks for never teasing me about leaving Montana to study marine mammals, and instead coming to visit and then moving here too. Last, but certainly not least, endless thanks to my actual family. Mom and Dad, I might still be bartending if you didn t constantly encourage me to follow my dream. Thanks for always dogsitting, helping me move too many times to count, and for keeping me on the family cell phone account. Your love and support has always been unwavering. Matt and Anthony thanks for believing in your nerdy sister, and for the fun times each trip home, often making me question coming back. Christina, you are the very best friend I could ask for, thank you for always checking on me to make sure I didn t die. Mike you and I both know I would have never made it this far without you. Thank you for reminding me how to enjoy the sunshine, that long walks cure all, and that sometimes I need to just stop working and watch 30 Rock. And Piper, of course, thank you for being my sidekick for this whole journey, forcing me to get some exercise, and covering everything I own in perfect little orange hairs.
8 TABLE OF CONTENTS Page CHAPTER 1: GENERAL INTRODUCTION... 1 Importance of sound to marine life... 1 Sources of increased anthropogenic noise... 2 Industrial sources... 2 Recreational sources... 3 Military sources... 3 Effects of noise on marine mammals... 3 Current regulations and the call for additional research... 4 Understanding the effects of noise: Behavioral response studies... 6 Current methods... 7 Limitations... 8 My research approach... 9 Northern elephant seals (Mirounga angustirostris) Life history Dive behavior Value as a study species References Figures CHAPTER 2: DEVELOPMENT AND EVALUATION OF AN ANIMAL- BORNE ACTIVE ACOUSTIC TAG TO CONDUCT MINIMALLY INVASIVE BEHAVIORAL RESPONSE STUDIES ON MARINE MAMMALS Abstract Introduction Results Novel tag development Deployments and basic dive behavior Controlled exposure experiment... 31
9 TABLE OF CONTENTS (Continued) Page Discussion Materials and methods Tag development Field efforts Behavioral analysis List of symbols and abbreviations Acknowledgements Author contributions Competing interests Funding References Figure legends Tables CHAPTER 3: GENERAL CONCLUSIONS AND FUTURE DIRECTIONS Primary findings Future directions Contribution to the field THESIS REFERENCES... 76
10 LIST OF FIGURES Figure Page Figure 1.1. Population Consequences of Acoustic Disturbance (PCAD) model Printed with permission from the National Research Council, Figure 2.1. Block diagram of prototype tag...52 Figure 2.2 Study site map with GPS track of all seal deployments...53 Figure 2.3 Study site map with GPS track of seal G Figure 2.4 Study site map with GPS track of seal G Figure 2.5 Study site map with GPS track of seal G Figure 2.6 Study site map with GPS track of seal G Figure 2.7 Study site map with GPS track of seal G Figure 2.8 Depth profiles of full deployments for all seals...59 Figure 2.9 Depth profiles for each dive with an exposure during an ascent...60 Figure Normalized change in depth against normalized change in time for all ascending dive inversions...61 Figure Normalized change in depth versus elapsed time for all dive inversions, seal G Figure Descent rates before and after exposures during descents...63 Figure 2.13 Spectrogram of flow noise before and after exposure...64 Figure 2.14 Flow noise levels before and after exposure...65 Figure 2.15 Spectrograms of playback tracks...66 Figure 2.16 Example of corrected and uncorrected depth profiles...67
11 LIST OF TABLES Table Page Table 2.1. Deployment details and dive statistics for all study individuals.68 Table 2.2 Number of exposures by seal and dive state..69 Table 2.3 Changes in depth for exposures and non-exposure dive inversions.70 Table 2.4 Comparison of number of responses as predicted, by exposure type 71 Table 2.5 Calibrated source levels for sound exposure tracks...72
12 Development and evaluation of an animal-borne active acoustic tag to conduct minimally invasive behavioral response studies on marine mammals CHAPTER 1: GENERAL INTRODUCTION Importance of sound to marine life Light attenuates quickly in seawater, so much of the ocean is dark beneath 200 m depth. Conversely, sound travels well underwater about five times as fast (~1,500 m/s) as it does in air (~330 m/s) and in the case of low frequency sounds, over very long distances (tens to hundreds of kilometers). Many marine animals have developed specialized acoustic sensory systems to perform basic life functions. Larval fish and crustaceans are known to cue in to the sound of coral reefs to navigate to recruitment locations (Tolimieri et al., 2000; Montgomery et al., 2006). Many fish species produce sounds to defend territories, attract mates, and maintain shoal cohesion (Amorim, 2006). Marine mammals, in particular, are well adapted to use sound in the ocean, and rely on sound for foraging, communication, navigation and migration, reproduction, and predator avoidance (Richardson et al., 1995). Marine mammals have particularly sensitive underwater hearing and odontocetes use echolocation to detect and differentiate between objects in their environment (Ketten, 2004; Au, 2008). Many marine mammals use sound in social contexts to find mates, communicate with offspring, or maintain family groups (Clapham and Mattila, 1990; Ford, 1991; Sayigh et al., 1998). Some species of baleen whales can even communicate over very long distances; under idea propagation conditions even several hundreds of kilometers (Cummings and Thompson, 1971; Payne and Webb, 1971; Croll et al., 2002). Because of the importance of sound to marine life, changes in ambient, or background, ocean noise conditions could have profound implications for species success and survival.
13 2 Sources of increased anthropogenic noise Ambient noise in parts of the world s oceans has increased over the last 50 years, in some areas exceeding 10 db re 1 µpa (Andrew et al., 2002; Hildebrand, 2005; Hildebrand, 2009). This rise in ambient noise in some of these ocean areas is associated with increased human industrial, recreational, and military use of the oceans. Industrial sources Commercial shipping is the largest contributor of anthropogenic noise, particularly at low-frequencies (<1 khz). Global shipping traffic increased fourfold in 20 years, from 1992 to 2012 (Tournadre, 2014). There are not only more ships, but they are larger, and traveling faster, which all translates to higher noise levels. For example, when shipping along the Eastern Seaboard of the United States and Canada shut down following September 11, 2001, ocean noise levels temporarily decreased by 6 db re 1 µpa (Rolland et al., 2012). Seismic surveys for petroleum reservoirs are another significant anthropogenic noise source and the number of surveys is likely to increase - especially in the Arctic Ocean as ice recedes and northern latitudes become more accessible by ship. These surveys use large, ship-towed, airgun arrays to generate high energy pressure waves which penetrate the water column and seafloor. Hydrophones, also towed behind the vessel, capture the signals reflected by the various substrate layers which provide information on oil and gas deposits. Arrays (tens of airguns) can produce source levels up to 259 db re 1 µpa and are usually operated for weeks at a time, with blasts occurring every 6-10 seconds (Richardson et al., 1995; Nieukirk et al., 2004). Construction of drilling platforms, wind and wave energy platforms, and oceanic mining can further increase ambient noise levels (Richardson et al., 1995; Bailey et al., 2010). Commercial fishing fleets also contribute significantly to ocean noise during heavy fishing periods (Richardson et al., 1995). All of these low-frequency inputs of noise overlap with the frequency range biologically relevant to baleen whales and fish.
14 3 Recreational sources Recreational sources of noise range in size from small jet propelled personal watercraft to mid-size whale watching or recreational fishing vessels up to thousand passenger cruise ships. These inputs have a higher contribution to ocean noise in coastal areas. Even these small vessels, although largely unregulated, can contribute substantially to local ambient noise, especially in the mid- to high-frequency ranges at which toothed whales echolocate and vocalize (Jensen et al., 2009). Military sources Military sources of noise include ship or aircraft engines and propellers and sonar operation. Low- and mid-frequency active sonar is used to scan large areas for enemy watercraft such as submarines. Sonar is used throughout the world s oceans for war-time operations and peace-time training, including joint training of maritime forces from allied countries. Military sources of noise are more intermittent than commercial inputs, but the sound pressure levels (240 db re 1 µpa) and tonal frequency characteristics of sonar pulses (similarities to killer whales, a known predator of marine mammals) may make sonar especially harmful to marine mammals (Jepson et al., 2003; Cox et al., 2006; D Amico et al., 2009). Effects of noise on marine mammals There is a variety of evidence that increased ambient noise levels and intermittent sound exposures to specific sounds have a measureable effect on marine mammals. Observed impacts range in severity from brief interruptions of basic life functions to physiological changes, acute injury and even death. The threat of acoustic masking and the ability for anthropogenic noise, such as that produced by shipping, to limit communication ranges is a conservation concern for many marine mammals species (Clark et al., 2009). Evidence exists that foraging by beaked whales may be interrupted by increased ship noise (Aguilar Soto et al., 2006) and foraging by sperm whales may decrease during seismic airgun activity (Miller et al., 2009). Gray whales altered their migration route in the Eastern North Pacific in response to simulated lowfrequency sonar (Tyack and Clark, 1998) and bowhead whales avoided normal
15 4 migration routes during seismic airgun operations in the Beaufort Sea (Richardson et al., 1999). Studies have also shown gray whales can be displaced from primary feeding and breeding grounds in response to increased noise from seismic surveys and ship playbacks (Jones et al., 1994; Weller et al., 2006). Rolland et al. (2012) found that decreased stress hormone levels in North Atlantic right whales coincided with a significant decrease in low-frequency ambient noise resulting from limited shipping after September 11, When shipping and noise levels increased again, stress hormone levels returned to pre-september 11 levels. Cetacean strandings have been connected to naval sonar and seismic airgun activity, and these strandings provide evidence for physical injury and some have resulted in mass mortalities (Jepson et al., 2003; Taylor et al., 2004; Cox et al., 2006; D Amico et al., 2009). In the last few decades, investigating specific impacts of noise on marine life has become a focus in marine mammal and conservation science (Southall et al., 2007; Popper and Hawkins, 2012). The potential for cumulative effects of even short duration responses may have profound implications for individual health, foraging and reproductive success, and population trends. However, acute and chronic behavioral and physiological changes in response to specific sounds are difficult to study experimentally, and therefore are still poorly understood. Current regulations and the call for additional research In the United States, the Marine Mammal Protection Act (1972), Endangered Species Act (1973), and National Environmental Policy Act (1969) require that appropriate regulations exist to protect marine mammals from human exploitation or disturbance, including acoustic harassment. Harassment is defined as: any act of pursuit, torment, or annoyance which (Level A Harassment) has the potential to injure a marine mammal or marine mammal stock in the wild; or (Level B Harassment) has the potential to disturb a marine mammal or marine mammal stock in the wild by causing disruption of behavioral patterns, including, but not limited to, migration, breathing, nursing, breeding, feeding, or sheltering but which does not have the potential to injure a marine mammal
16 5 or marine mammal stock in the wild (1994 Amendments to the MMPA, source: NOAA). The National Marine Fisheries Service (NMFS) and US Fish and Wildlife Service (FWS) are the governing bodies responsible for issuing permits for allowable instances of marine mammal harassment, either for scientific research, marine industrial activities, or military exercises. Based on current data, underwater sound levels which constitute Level A Harassment are those that exceed 190 dbrms re 1 µpa for pinnipeds and 180 dbrms re 1 µpa for cetaceans. Impulsive sounds exceeding 160 dbrms re 1 µpa and non-impulsive sounds exceeding 120 dbrms re 1 µpa constitute Level B Harassment, for both pinnipeds and cetaceans 1. Currently, these criteria group together all cetacean species and pinniped species, regardless of their hearing abilities or scientific knowledge of normal behavior, but proposed changes have been outlined (NOAA, 2013). In order for NMFS and FWS to create and enforce appropriate regulations, sufficient data on how anthropogenic noise affects marine mammals is needed. Because of the current paucity of data, these regulations may be inappropriate or insufficient (Gilchrest et al., 2008; NOAA, 2013; Tougaard et al., 2014). The National Research Council (NRC) created the Population Consequences of Acoustic Disturbance (PCAD) model (Figure 1.1) to serve as a conceptual framework that outlines the path from initial acoustic stimulus to population level effects of exposure to that stimulus (National Research Council, 2005). Additionally, the model identifies specific research needs and the ability of current methods to answer those research needs. The PCAD model contains five levels, each consisting of a group of related variables of interest. These five levels are arranged in order of increasing scope: (1) characteristics of the sound, (2) changes in behavior, (3) effects on life functions, (4) changes in vital rates, and (5) population level effects. The relationship connecting sequential levels is represented by one of four transfer functions. Each level is scored by how well the group of variables can be measured, and each transfer function is scored by how well the relationship between variables is currently understood (Figure 1.1, boxes and arrows, respectively). Characterizing the 1 Source: NOAA. threshold_guidance.html
17 6 actual sound of interest frequency characteristics, loudness, and duration is relatively straightforward (first level). Examining direct changes in behavior of individuals is more difficult (second level). Investigating life functions affected by changes in behavior (third level), potential vital rate changes (fourth level), and resulting population effects (fifth level) is very difficult in marine mammals because of their mostly or entirely aquatic lifestyle. Some data exists relating acoustic stimuli with direct changes in behavior of individuals (first transfer function), and connecting those specific changes in behavior to immediate effects on life functions (second transfer function). There are little to no data relating effects on life functions to long-term changes in vital rates (third transfer function, symbolized by 0 ). The connection between vital rates and population level effects (fourth transfer function) is relatively well understood despite the difficulty to study each of those variables independently. The NRC (2005) identified the third transfer function (Figure 1.1, arrow 3), the influence of interrupted life functions on vital rates, as a gap in knowledge and data, and thus an area of high research need and importance. Yet, not many viable, effective methodologies currently exist that allow investigation of this relationship between behavioral changes in life function due to noise and the subsequent effect on vital rates. Many viable research approaches are constrained by cost or limitations in sample size. Present studies either take place in captive facilities, or are only conducted over very short time periods, limiting the ability of researchers to understand effects on freeranging individuals and over large time scales. Development of new research tools is needed to fully understand the potential effects of a noisier ocean on individual and populations of marine mammals (Tyack, 2009). Understanding the effects of noise: Behavioral response studies One of the most important research techniques for studying the effects of anthropogenic noise on marine mammals is a behavioral response study (BRS), which is a unique type of playback experiment concerned with changes in a target animals behavior to a highly controlled, measured acoustic stimulus. BRS provides an effective method to study discrete behavioral responses of individual, free-ranging marine mammals to particular sounds, and data collected are used to inform regulatory
18 agencies of the potential negative effects of anthropogenic sound sources on the animals (Tyack, 2009; Southall et al., 2012). 7 Current methods Acoustic playback studies use motion sensing tags, visual observations, or physiological measurements to monitor potential responses of animals to particular sounds. They are conducted to help understand how individual animals use sounds in social contexts, as foraging cues, and to avoid predators or how hormone levels may change in response to loud sound exposures (e.g. Charrier et al., 2002; Hayes et al., 2004; Romano et al., 2004; Curé et al., 2012; Curé et al., 2013); they have even been used to coarsely study hearing frequency ranges (Ghoul and Reichmuth, 2014). A controlled exposure experiments (CEE) is a playback experiment with additional focus on specific acoustic properties of the playback stimulus (e.g. loudness, frequency range) and how that influences an individual s level of response, either physical or behavioral (Tyack, 2009). In the last decade BRS have emerged as a specific type of acoustic playback, using a CEE design, to study discrete behavioral responses of freeranging whales to simulated mid-frequency active sonar (Tyack et al., 2011; Miller et al., 2012; Southall et al., 2012; DeRuiter et al., 2013; Goldbogen et al., 2013; Stimpert et al., 2014) as well as shipping noise (Aguilar Soto et al., 2006) and seismic airgun activity (Madsen et al., 2006; Jochens et al., 2008). These studies directly inform regulatory agencies of the negative effects of anthropogenic sound sources on animal behavior, influencing future management decisions regarding use of sonar by the US Navy (training areas and protocol), designation of shipping lanes and speed regulations, and time and cycle maximums for seismic activity. Typically, BRS involve tagging an animal with a passive acoustic recorder and archival motion sensors (e.g., DTAG and Acousonde; Johnson and Tyack, 2003 and then using a ship-mounted sound source to project a high source level sound towards the animal, monitoring the animal s behavior before, during, and after exposure. Locomotive behavior is recorded using a 3-axis accelerometer and magnetometer, dive behavior and depth is recorded using a pressure sensor, and an acoustic recorder allows the actual sound level the animal received to be measured, as
19 8 well as detection of potential changes in vocal behavior. The archival tag is released 2-24 hours after attachment and recovered. In situations where tags are not available, visual observations are made from a small boat before, during, and after exposure. The stimulus has a controlled source level and duration. Observations of the animals behavior before and after the exposure are used as a baseline to behavior during the exposure period. Limitations BRS have been instrumental in observing and quantifying responses of marine mammals to specific acoustic stimuli, however, current studies are limited financially, logistically, and in the data collected. Under the current configuration, findings from behavioral response studies do not address the third transfer function of the PCAD model, connecting alteration of life functions to long-term changes in vital rates (Figure 1.1). Traditionally, BRS are expensive to conduct; they require multiple vessels and the necessary personnel to man small tagging boats, conduct visual observations from a large ship, manage the sound source, and other teams. Current suction cup tags used provide fine-scale behavioral data, but are difficult to attach and operate on a very restricted time scale they rarely stay on for over 24 hours often resulting in very small sample sizes and limited exposures and baseline behavior per animal. However, researchers are working on overcoming these issues within the basic external sound source experimental design (Southall et al., 2012). Some limitations of the current BRS approach cannot be addressed within the basic off-animal sound source approach. Because the sound is played from a ship some distance from the individual, and both the ship and animal are moving independently, actual sound level the animal receives cannot be well controlled during the playback. However, estimates can be calculated using propagation modelling before the playback begins and the final received level is recorded on the acoustic tag and retrieved with tag recovery. These experiments expose the study individual to a high source level sound (up to ~210 db re 1 µpa; Southall et al., 2012) for the value of collecting necessary data, however, there is risk to other animals in the area and they may be subsequently exposed, with no valuable behavioral data collected. Permitting
20 9 restrictions require experiments to be halted if non-study animals enter the at-risk area which means sometimes time and resources are spent to collect incomplete, unusable data. Finally, these studies are most often conducted on or near Navy training ranges, both because animals in those areas are directly at risk, and because the ranges provide bottom-moored hydrophone arrays which can help locate elusive species and increase tagging success. However, by conducting experiments on animals that are known to frequent areas with high occurrence of sonar, there is a possibility that animals being tagged and exposed are habituated to such sounds and will not react to the same degree as a non-habituated animal. The current BRS approach has been valuable in collecting data on individual responses to particular sounds that result in immediate changes in behavior (the first transfer function) and interruption or alternation of basic life functions (second transfer function), but are limited from looking at long-term effects of exposure or effects of long-term, repeated exposures and habituation. If scientists are going to successfully address the third transfer function of the PCAD model, additional methodologies must be developed (Tyack et al., 2003; Southall et al., 2009). My research approach Combining the playback sound source with the behavioral sensors and acoustic recorder into one animal-mounted tag could potentially allow for an alternative BRS method that overcomes some of the limitations of the current configuration. Such a tag would allow for better control of exposure sound levels, maximum exposure level for the target animal with minimum exposure to nearby individuals, and long-term deployments, enabling investigation of the effects of varied sound levels, repeated exposure, and habituation, all in a more cost effective package. I worked with others to coordinate the development, integration, and testing of a prototype acoustic tag to test its effectiveness as an alternative method to conduct behavioral response studies. Field testing of this new technology was conducted using a well-studied, easily accessible animal, the northern elephant seal. This provided the opportunity for high success finding target individuals, relative ease in attaching and deploying tags, and reliable return of study individuals for tag recovery, making
21 10 evaluation of the prototype tag an achievable main study objective and outcome. Results of this effort and proof of concept for this tag could expand current data regarding behavioral responses of marine mammals to anthropogenic noise and provide enhanced understanding of long-term effects, leading to better informed regulation of human-generated sound. Northern elephant seals (Mirounga angustirostris) Life history Northern elephant seals, Mirounga angustirostris (Gill 1866), are a large, deepdiving phocid species with an uncommon life history (Stewart et al., 1994). They were hunted nearly to extinction in the late 19 th century, but have since recovered from just a few individuals at Isla de Guadalupe to almost 170,000 individuals at 16 breeding colonies from Natividad in Baja California ( N, W) to Pt. Reyes, California (37 59' 48" N, ' 11 W; Le Boeuf et al., 2011). Northern elephant seals are a deep diving pinniped, with both sexes diving to mean depths of 500 m and maximum depths of up to 1735 m, and they spend the majority of their lives at sea (Le Boeuf et al., 1988; DeLong and Stewart, 1991; Le Boeuf, 1994; Robinson et al., 2012). Seals haul out at discreet breeding colonies two times a year, to breed within a polygynous system and to undergo a catastrophic molt where their entire skin and coat is replaced over a few weeks and they cannot go into the water (Worthy et al., 1992). Different sex and age classes appear at the beach to molt at different times of the summer and fall, and adult females and males both return during the winter months to give birth and mate, respectively (Worthy et al., 1992; Stewart et al., 1994). When not hauled out, the seals embark on 2-3 month foraging trips traveling thousands of kilometers to the central north Pacific or Aleutian Islands to feed on squid and demersal fish (Stewart and DeLong, 1994). Northern elephant seals exhibit high site fidelity and often return to where they were born year after year (Le Boeuf et al., 2011). Killer whales and white sharks are the only known predators of northern elephant seals (Le Boeuf and Crocker, 1996). Elephant seals use vocal signaling on land during the breeding season to defend themselves, their pups, or their harem, but are largely silent and solitary while on foraging trips at sea (Le Boeuf and Petrinovich, 1974; Burgess et
22 11 al., 1998). Northern elephant seals have the widest range and most sensitive underwater hearing of any phocid measured, with higher sensitivities at lower frequencies (72.8 db re 1 µpa at 200 Hz to 59 db re 1 µpa at 6300 Hz) than other pinnipeds (Kastak and Schusterman, 1998; Kastak and Schusterman, 1999; Reichmuth et al., 2013). Dive behavior Northern elephant seals exhibit stereotypic dive behavior with up to 90% of their time in the open ocean spent performing repeated deep dives to typical depths of over 300 m for adult males and over 500 m for adult females and maximum depths of over 1500 m (adult male) and over 1700 m (adult female; Le Boeuf, 1994; Robinson et al., 2012). Dive durations are typically about 20 min, with durations up to 109 minutes recorded in an adult female (Le Boeuf et al., 2000; Robinson et al., 2012). As seals move offshore, dives appear to follow the seafloor until reaching the continental shelf (Le Boeuf et al., 1988; Le Boeuf et al., 1993). Diving patterns have been well studied since the first time depth recorders (TDRs) were created to the fine-scale 3D motion sensors combined with TDRs that are available today (Le Boeuf, 1994; Robinson et al., 2012). A translocation study found juvenile seals use continuous stroking until reaching about 20 m depth, then switch to stroke-and-glide swimming until 60 m, after which they used prolonged gliding in a passive descent (Davis et al., 2001). The ascent is marked by mostly continuous stroking (Davis et al., 2001). Although it is likely the continuous deep diving exhibited by northern elephant seals is related to foraging strategies and distribution or energy conservation, it has been hypothesized that diving deep also serves as a predator avoidance response (Le Boeuf et al., 1988). They can dive significantly deeper than their known predators, spend 60% of their day out of depth range of those predators, and adults spend little time on the shallow continental shelf (Le Boeuf et al., 1988; Le Boeuf and Crocker, 1996). Resident killer whales, which only feed on fish but have been more well studied, have maximum dive depths averaging ± 61.8 m but spend 66% of their time above 20 m while transient killer whales, which feed on marine mammals, spend most of their time between 20 and 60 m (Baird, 1994; Baird, 2000; Baird et al., 2005). White
23 sharks depth range east of 125 W in North Pacific, the area of their range in which they feed on pinnipeds, is m (Boustany et al., 2002). 12 Value as a study species Northern elephant seals have been studied at Año Nuevo State Park in Northern California for over 50 years, and are an ideal study species for testing new tag technologies and BRS methodologies. Juvenile (1-2 year old) seals have been shown to carry instruments up to 5.6 kg (up to 3.8% of body weight) without altering diving and swimming behavior (Le Boeuf et al., 1986; Fletcher et al., 1996) and are easily approachable on the beach. The catastrophic molt allows minimally invasive attachment techniques that stay on for long periods of time. Quick drying epoxy is used to glue the instruments on, and the glue falls off with the molting fur (Le Boeuf et al., 1988). Because of high site fidelity, most juvenile seals transported away from the beach will reliably return within a few days or weeks, and adult seals instrumented before spring foraging trips will likely return to the haul out site they departed from. Previous studies found 88% of juvenile seals transported and released away from Año Nuevo returned to the beach within 21 days, and females instrumented after weaning their pups returned after 72.6 ± 5.0 days (Le Boeuf et al., 1988; Oliver et al., 1998). Diving performance during translocations is similar to dive performance during migrations (Le Boeuf et al., 1996). Additionally, northern elephant seals are a marine mammal of concern with respect to anthropogenic noise (Kastak and Schusterman, 1999). They frequently dive as deep as or deeper than the deep sound channel, where low frequency sounds can travel long distances due to a sound speed minimum created by changes in temperature, pressure, and salinity in the water column (Le Boeuf et al., 1988; Payne and Webb, 1971). This sound speed minimum causes low frequency sound waves to refract up and down through one horizontal channel, allowing low-frequency sound waves to travel thousands of kilometers without the signal losing significant energy. In mid-latitudes, this channel is located between 600 and 1000 m. Northern elephant seals breed and haul out on locations near high human use areas, including areas of naval sonar training, and they have the best underwater hearing sensitivity at the lowest frequencies of all
24 phocids that have been tested, perhaps putting them at higher risk to increased manmade noise (Kastak and Schusterman, 1998; Kastak and Schusterman, 1999). 13
25 14 References Aguilar Soto, N., Johnson, M., Madsen, P. T., Tyack, P. L., Bocconcelli, A. and Fabrizio Borsani, J. (2006). Does intense ship noise disrupt foraging in deepdiving cuvier s beaked whales (Ziphius Cavirostris)? Mar. Mammal Sci. 22, Amorim, M. C. P. (2006). Diversity of Sound Production in Fish. In Communication in Fishes (ed. Ladich, F., Collin, S. P., Moller, P., and Kappor, B. G.), pp Enfield, NH: Science Publishers. Andrew, R. K., Howe, B. M., Mercer, J. A. and Dzieciuch, M. A. (2002). Ocean ambient sound: Comparing the 1960s with the 1990s for a receiver off the California coast. Acoust. Res. Lett. Online 3, Au, W. W. L. (2008). Echolocation. In Encyclopedia of Marine Mammals (ed. Perrin, W. F., Wursig, B., and Thewissen, J. G. M.), pp San Diego: Academic Press. Bailey, H., Senior, B., Simmons, D., Rusin, J., Picken, G. and Thompson, P. M. (2010). Assessing underwater noise levels during pile-driving at an offshore windfarm and its potential effects on marine mammals. Mar. Pollut. Bull. 60, Baird, R. W. (1994). Foraging behaviour and ecology of transient killer whales (Orcinus orca). PhD dissertation, Simon Fraser University, Burnaby, BC, Canada. Baird, R. W. (2000). The Killer Whale. Cetacean Soc Baird, R. W., Hanson, M. B. and Dill, L. M. (2005). Factors influencing the diving behaviour of fish-eating killer whales: sex differences and diel and interannual variation in diving rates. Can. J. Zool. 83, Boustany, A. M., Davis, S. F., Pyle, P., Anderson, S. D., Le Boeuf, B. J. and Block, B. A. (2002). Expanded niche for white sharks. Nature 415, Burgess, W. C., Tyack, P. L., Le Boeuf, B. J. and Costa, D. P. (1998). A programmable acoustic recording tag and first results from free-ranging northern elephant seals. Deep. Res. Part II Top. Stud. Oceanogr. 45,
26 15 Charrier, I., Mathevon, N., Hassnaoui, M., Carraro, L. and Jouventin, P. (2002). The subantarctic fur seal pup switches its begging behaviour during maternal absence. Can. J. Zool. 80, Clapham, P. J. and Mattila, D. K. (1990). Humpback whale songs as indicators of migration routes. Mar. Mammal Sci. 6, Clark, C., Ellison, W., Southall, B., Hatch, L., Van Parijs, S., Frankel, a and Ponirakis, D. (2009). Acoustic masking in marine ecosystems: intuitions, analysis, and implication. Mar. Ecol. Prog. Ser. 395, Cox, T., Ragen, T., Read, A., Vos, E., Baird, R. W., Balcomb, K., Barlow, J., Caldwell, J., Cranford, T., Crum, L., et al. (2006). Understanding the impacts of anthropogenic sound on beaked whales. J. Cetacean Res. Manag. 7, Croll, D. A., Clark, C. W., Acevedo, A., Tershy, B. R., Flores, S., Gedamke, J. and Urban, J. (2002). Bioacoustics: Only male fin whales sing loud songs. Nature 417, Cummings, W. C. and Thompson, P. O. (1971). Underwater sounds from the blue whale, Balaenoptera musculus. J. Acoust. Soc. Am. 50, Curé, C., Antunes, R., Samarra, F., Alves, A. C., Visser, F., Kvadsheim, P. H. and Miller, P. J. O. (2012). Pilot whales attracted to killer whale sounds: acoustically-mediated interspecific interactions in cetaceans. PLoS One 7, e Curé, C., Antunes, R., Alves, A. C., Visser, F., Kvadsheim, P. H. and Miller, P. J. O. (2013). Responses of male sperm whales (Physeter macrocephalus) to killer whale sounds: implications for anti-predator strategies. Sci. Rep. 3, D Amico, A., Gisiner, R. C., Ketten, D. R., Hammock, J. a., Johnson, C., Tyack, P. L. and Mead, J. (2009). Beaked whale strandings and naval exercises. Aquat. Mamm. 35, Davis, R. W., Fuiman, L. A., Williams, T. M. and Le Boeuf, B. J. (2001). Threedimensional movements and swimming activity of a northern elephant seal. Comp. Biochem. Physiol. Part A 129, DeLong, R. L. and Stewart, B. S. (1991). Diving patterns of northern elephant seal bulls. Mar. Mammal Sci. 7,
27 16 DeRuiter, S. L., Southall, B. L., Calambokidis, J., Zimmer, W. M. X., Sadykova, D., Falcone, E. A., Friedlaender, A. S., Joseph, J. E., Moretti, D., Schorr, G. S., et al. (2013). First direct measurements of behavioural responses by Cuvier s beaked whales to mid-frequency active sonar. Biol. Lett. 9, Fletcher, S., Le Boeuf, B. J., Costa, D. P., Tyack, P. L. and Blackwell, S. B. (1996). Onboard acoustic recording from diving northern elephant seals. J. Acoust. Soc. Am. 100, Ford, J. K. B. (1991). Vocal traditions among resident killer whales (Orcinus orca) in coastal waters of British Columbia. Can. J. Zool. 69, Ghoul, A. and Reichmuth, C. (2014). Hearing in sea otters (Enhydra lutris): Audible frequencies determined from a controlled exposure approach. Aquat. Mamm. 40, Gilchrest, Y. V, Fetherston, T. N. and Neales, B. E. (2008). Assessing potential sites for undersea training ranges: the effects of active sonars on marine mammals. NUWC-NPT Tech. Rep. 11, Goldbogen, J. A., Southall, B. L., Deruiter, S. L., Calambokidis, J., Friedlaender, A. S., Hazen, E. L., Falcone, E. A., Schorr, G. S., Douglas, A., Moretti, D., et al. (2013). Blue whales respond to simulated mid-frequency military sonar. Proc. R. Soc. B Biol. Sci. 280, Hayes, S. A., Kumar, A., Costa, D. P., Mellinger, D. K., Harvey, J. T., Southall, B. L. and Le Boeuf, B. J. (2004). Evaluating the function of the male harbour seal, Phoca vitulina, roar through playback experiments. Anim. Behav. 67, Hildebrand, J. A. (2005). Impacts of Anthropogenic Sound. In Marine Mammal Research: Conservation Beyond Crisis (ed. Reynolds, III, J. E., Perrin, W. F., Reeves, R. R., Montgomery, S., and Ragen, T. J.), pp Baltimore, Maryland: The John Hopkins University Press. Hildebrand, J. (2009). Anthropogenic and natural sources of ambient noise in the ocean. Mar. Ecol. Prog. Ser. 395, 5 20.
28 17 Jensen, F., Bejder, L., Wahlberg, M., Aguilar de Soto, N., Johnson, M. and Madsen, P. (2009). Vessel noise effects on delphinid communication. Mar. Ecol. Prog. Ser. 395, Jepson, P. D., Arbelo, M., Deaville, R., Patterson, I. A. P., Castro, P., Baker, J. R., Degollada, E., Ross, H. M., Herráez, P., Pocknell, A. M., et al. (2003). Gas-bubble lesions in stranded cetaceans. Nature 425, Jochens, A., Biggs, D., Benoit-Bird, K., Engelhardt, D., Gordon, J., Hu, C., Jaquet, N., Johnson, M., Leben, R., Mate, B., et al. (2008). Sperm Whale Seismic Study in the Gulf of Mexico: Synthesis Report. U.S. Dep. Inter. Miner. Manag. Serv. Gulf Mex. OCS Reg. New Orleans, LA. OCS Study MMS Johnson, M. P. and Tyack, P. L. (2003). A digital acoustic recording tag for measuring the response of wild marine mammals to sound. IEEE J. Ocean. Eng. 28, Jones, M., Swartz, S. and Dahlheim, M. (1994). Census of gray whale abundance in San Ignacio Lagoon: a follow-up study in response to low whale counts recorded during an acoustic playback study. Rep. No. NTIS PB to US Mar. Mammal Comm. Kastak, D. and Schusterman, R. J. (1998). Low-frequency amphibious hearing in pinnipeds: methods, measurements, noise, and ecology. J. Acoust. Soc. Am. 103, Kastak, D. and Schusterman, R. J. (1999). In-air and underwater hearing sensitivity of a northern elephant seal (Mirounga angustirostris). Can. J. Zool. 77, Ketten, D. R. (2004). Marine mammal auditory systems : A summary of audiometric and anatomical data and implications for underwater acoustic impacts. Polarforschung 72, Le Boeuf, B. J. (1994). Variation in the Diving Pattern of Northern Elephant Seals with Age, Mass, Sex, and Reproductive Condition. In Elephant Seals: Population Ecology, Behavior, and Physiology (ed. Le Boeuf, B. J. and Laws, R. M.), pp Berkeley: University of California Press.
29 18 Le Boeuf, B. J. and Crocker, D. E. (1996). Diving Behavior of Elephant Seals: Implications for Predator Avoidance. In Great White Sharks: The Biology of Carcharodon carcharias (ed. Klimley, A. P. and Ainley, D. G.), pp San Diego: Academic Press. Le Boeuf, B. J. and Petrinovich, L. F. (1974). Dialects of northern elephant seals, Mirounga angustirostris: Origin and reliability. Anim. Behav. 22, Le Boeuf, B. J., Costa, D. P., Huntley, A. C., Kooyman, G. L. and Davis, R. W. (1986). Pattern and depth of dives in northern elephant seals, Mirounga angustirostris. J. Zool. 208, 1 7. Le Boeuf, B. J., Costa, D. P., Huntley, A. C. and Feldkamp, S. D. (1988). Continuous, deep diving in female northern elephant seals, Mirounga angustirostris. Can. J. Zool. 66, Le Boeuf, B. J., Crocker, D. E., Blackwell, S. B., Morris, P. A. and Thorson, P. H. (1993). Sex differences in diving and foraging behaviour of northern elephant seals. Symp. Zool. Soc. London 66, Le Boeuf, B. J., Morris, P. A., Blackwell, S. B., Crocker, D. E. and Costa, D. P. (1996). Diving behavior of juvenile northern elephant seals. Can. J. Zool. 74, Le Boeuf, B. J., Crocker, D. E., Costa, D. P., Blackwell, S. B., Webb, P. M. and Houser, D. S. (2000). Foraging ecology of northern elephant seals. Ecol. Monogr. 70, Le Boeuf, B. J., Condit, R., Morris, P. A. and Reiter, J. (2011). The northern elephant seal (Mirounga angustirostris) rookery at Año Nuevo: A case study in colonization. Aquat. Mamm. 37, Madsen, P. T., Johnson, M., Miller, P. J. O., Aguilar Soto, N., Lynch, J. and Tyack, P. (2006). Quantitative measures of air-gun pulses recorded on sperm whales (Physeter macrocephalus) using acoustic tags during controlled exposure experiments. J. Acoust. Soc. Am. 120, Miller, P. J. O., Johnson, M. P., Madsen, P. T., Biassoni, N., Quero, M. and Tyack, P. L. (2009). Using at-sea experiments to study the effects of airguns on
30 19 the foraging behavior of sperm whales in the Gulf of Mexico. Deep Sea Res. Part I Oceanogr. Res. Pap. 56, Miller, P. J. O., Kvadsheim, P. H., Lam, F. P. a, Wensveen, P. J., Antunes, R., Alves, A. C., Visser, F., Kleivane, L., Tyack, P. L. and Sivle, L. D. (2012). The Severity of behavioral changes observed during experimental exposures of killer (Orcinus orca), long-finned Pilot (Globicephala melas), and sperm (Physeter macrocephalus) whales to naval sonar. Aquat. Mamm. 38, Montgomery, J. C., Jeffs, A., Simpson, S. D., Meekan, M. and Tindle, C. (2006). Sound as an orientation cue for the pelagic larvae of reef fishes and decapod crustaceans. Adv. Mar. Biol. 51, National Research Council (2005). Marine mammal populations and ocean noise: determining when noise causes biologically significant effects. Washington, D.C., D.C.: U.S. National Academy of Sciences. Nieukirk, S. L., Stafford, K. M., Mellinger, D. K., Dziak, R. P. and Fox, C. G. (2004). Low-frequency whale and seismic airgun sounds recorded in the mid- Atlantic Ocean. J. Acoust. Soc. Am. 115, NOAA (2013). Draft Guidance for Assessing the Effects of Anthropogenic Sound on Marine Mammals - Acoustic Threshold Levels for Onset of Permanent and Temporary Threshold Shifts. Maryland. Oliver, G. W., Morris, P. A., Thorson, P. H. and Le Boeuf, B. J. (1998). Homing behavior of juvenile northern elephant seals. Mar. Mammal Sci. 14, Payne, R. and Webb, D. (1971). Orientation by means of long range acoustic signaling in baleen whales. Ann. N. Y. Acad. Sci. 188, Popper, A. and Hawkins, A. D. (2012). Effects of noise on aquatic life. New York: Springer. Reichmuth, C., Holt, M. M., Mulsow, J., Sills, J. M. and Southall, B. L. (2013). Comparative assessment of amphibious hearing in pinnipeds. J. Comp. Physiol. A Neuroethol. Sensory, Neural, Behav. Physiol. 199, Richardson, W. J., Greene, C. R., Malme, C. I. and Thomson, D. H. (1995). Marine Mammals and Noise. San Diego: Academic Press.
31 20 Richardson, W. J., Miller, G. W. and Greene, C. R. (1999). Displacement of migrating bowhead whales by sounds from seismic surveys in shallow waters of the Beaufort Sea. J. Acoust. Soc. Am. 106, Robinson, P. W., Costa, D. P., Crocker, D. E., Gallo-Reynoso, J. P., Champagne, C. D., Fowler, M. a., Goetsch, C., Goetz, K. T., Hassrick, J. L., Hückstädt, L. a., et al. (2012). Foraging behavior and success of a mesopelagic predator in the northeast Pacific Ocean: insights from a data-rich species, the northern elephant seal. PLoS One 7, e Rolland, R. M., Parks, S. E., Hunt, K. E., Castellote, M., Corkeron, P. J., Nowacek, D. P., Wasser, S. K. and Kraus, S. D. (2012). Evidence that ship noise increases stress in right whales. Proc. R. Soc. B Biol. Sci. 279, Romano, T. a, Keogh, M. J., Kelly, C., Feng, P., Berk, L., Schlundt, C. E., Carder, D. a and Finneran, J. J. (2004). Anthropogenic sound and marine mammal health: measures of the nervous and immune systems before and after intense sound exposure. Can. J. Fish. Aquat. Sci. 61, Sayigh, L. S., Tyack, P. L., Wells, R. S., Solows, A. R., Scott, M. D. and Irvine, A. B. (1998). Individual recognition in wild bottlenose dolphins: a field test using playback experiments. Anim. Behav Southall, B. L., Bowles, A. E., Ellison, W. T., Finneran, J. J., Gentry, R. L., Greene, C. R., Kastak, D., Ketten, D. R., Miller, J. H., Nachtigall, P. E., et al. (2007). Marine mammal noise exposure criteria: Initial scientific recommendations. Aquat. Mamm. 33, Southall, B. L., Berkson, J., Bowen, D., Brake, R., Eckman, J., Field, J., Gisner, R. C., Gregerson, S., Lang, W., Lewandowski, J., et al. (2009). Addressing the effects of human-generated sound on marine life: an integrated research plan for US federal agencies. Interagency Task Force on Anthropogenic Sound and the Marine Environment of the Joint Subcommittee on Ocean Science and Technology. Washington, D.C. Southall, B. L., Moretti, D., Abraham, B., Calambokidis, J., DeRuiter, S. L. and Tyack, P. L. (2012). Marine mammal behavioral response studies in Southern
32 21 California: Advances in technology and experimental methods. Mar. Technol. Soc. J. 46, Stewart, B. S. and DeLong, R. L. (1994). Postbreeding Foraging Migrations of Northern Elephant Seals. In Elephant Seals: Population Ecology, Behavior, and Physiology (ed. Le Boeuf, B. J. and Laws, R. M.), pp Berkeley: University of California Press. Stewart, B. S., Yochem, P. K., Huber, H. R., DeLong, R. L., Jameson, R. J., Sydeman, W. J., Allen, S. G. and Le Boeuf, B. J. (1994). History and present status of the northern elephant seal population. In Elephant Seals: Population Ecology, Behavior, and Physiology (ed. Le Boeuf, B. J. and Laws, R. M.), pp Berkeley: University of California Press. Stimpert, A. K., Deruiter, S. L., Southall, B. L., Moretti, D. J., Falcone, E. A., Goldbogen, J. A., Friedlaender, A., Schorr, G. S. and Calambokidis, J. (2014). Acoustic and foraging behavior of a tagged Baird s beaked whale (Berardius bairdii) exposed to simulated sonar. Sci. Rep. 4, 1 8. Taylor, B., Barlow, J., Pitman, R., Ballance, L., Klinger, T., DeMaster, D., Hildebrand, J., Urban, J., Palacios, D. and Mead, J. (2004). A call for research to assess risk of acoustic impact on beaked whale populations. Int. Whal. Comm. Sci. Comm. SC/56/E36,. Tolimieri, N., Jeffs, A. and Montgomery, J. C. (2000). Ambient sound as a cue for navigation by the pelagic larvae of reef fishes. Mar. Ecol. Prog. Ser. 207, Tougaard, J., Wright, A. J. and Madsen, P. T. (2014). Cetacean noise criteria revisited in the light of proposed exposure limits for harbour porpoises. Mar. Pollut. Bull. 90, Tournadre, J. (2014). Anthropogenic pressure on the open ocean: The growth of ship traffic revealed by altimeter data analysis. Geophys. Res. Lett. 41, Tyack, P. L. (2009). Acoustic playback experiments to study behavioral responses of free-ranging marine animals to anthropogenic sound. Mar. Ecol. Prog. Ser. 395,
33 22 Tyack, P. L. and Clark, C. W. (1998). Quick-look report: Playback of lowfrequency sound to gray whales migrating past the central California coast. Tyack, P. L., Gordon, J. and Thompson, D. (2003). Controlled exposure experiments to determine the effects of noise on marine mammals. Mar. Technol. Soc. J. 37, Tyack, P. L., Zimmer, W. M. X., Moretti, D., Southall, B. L., Claridge, D. E., Durban, J. W., Clark, C. W., D Amico, A., DiMarzio, N., Jarvis, S., et al. (2011). Beaked whales respond to simulated and actual navy sonar. PLoS One 6, e Weller, D. W., Rickards, S. H., Bradford, A. L., Burdin, A. M. and Brownell Jr, R. L. (2006). The influence of 1997 sesimic surveys on the behavior of western gray whales of Sakhalin Island, Russia. IWC Sci. Comm. SC/58/E4, 12. Worthy, G. A. J., Morris, P. A., Costa, D. P. and Le Boeuf, B. J. (1992). Moult energetics of the northern elephant seal (Mirounga angustirostris). J. Zool. Soc. London 227,
34 Figures 23 Figure 1.1. Reprinted with permission from National Research Council, 2005, Figure 3-1. Population Consequences of Acoustic Disturbance (PCAD) model. Each of the five levels are groups of variables of interest, connected by four transfer functions (numbered arrows). The "+" signs inside boxes indicate how well a group of variables can be measured and the + signs between boxes indicate how well the relationship between variables is understood, from 0 meaning unknown to +++ meaning easy to observe or well known.
35 24 CHAPTER 2: DEVELOPMENT AND EVALUATION OF AN ANIMAL- BORNE ACTIVE ACOUSTIC TAG TO CONDUCT MINIMALLY INVASIVE BEHAVIORAL RESPONSE STUDIES ON MARINE MAMMALS Selene Fregosi Intended for publication in the Journal of Experimental Biology with the following co-authors: Holger Klinck, Markus Horning, Daniel Costa, David Mann, Kenneth Sexton, Luis Huckstadt, David K. Mellinger, and Brandon L. Southall.
36 25 Abstract Development of new research tools is needed to better understand the potential effects of a noisier ocean on individual and populations of marine mammals. Current behavioral response studies utilize ship-mounted sound sources to induce short-term noise-related behavioral responses in tagged animals. Combining the tag with the sound source into an animal-mounted active acoustic and motion sensing tag could potentially allow for long-term, more controlled and cost-effective behavioral response studies. Field tests were conducted on juvenile northern elephant seals, Mirounga angustirostris, using both natural and anthropogenic stimuli, to evaluate the potential of such a prototype tag. Results showed that such an instrument does elicit behavioral responses in tagged individuals. Responses during the ascending phase of a dive consisted of a dive inversion, with the animal diving as deep as or deeper than its original dive depth (seven of nine exposures). Change in dive depth following exposure was significantly larger than change in depth for non-exposure inversions. A single exposure at the bottom phase of a deep dive followed the same pattern. Dive inversions were observed following white noise, sperm whale clicks, killer whale whistles, and sonar exposures, but not following common dolphin whistles. Responses to exposures received during the descending phase of a dive resulted in an increased descent rate in nine of ten exposures. All eight exposures during shallow dives, where the animals were likely limited by bathymetry from diving any deeper, were characterize by increased flow noise following exposure, an indicator of increased swim speed. Results showing differential responses to specific exposure stimuli were inconclusive. Tag improvements and additional field efforts are needed to validate the tag s use in behavioral response studies to specific acoustic stimuli. There is potential to use this technology to study physiological effects of extended deep dives on marine mammals as well as frequency dependent hearing because of its ability to induce prolonged, unplanned dives in response to man-made sounds.
37 26 Introduction For marine mammal species that spend most of their lives underwater, sound plays an important sensory role. Marine mammals rely on acoustic cues for many life functions including navigation, foraging, communication, and predator avoidance (Richardson et al., 1995). Ocean ambient noise levels have risen in certain areas due to increased anthropogenic activities (Hildebrand, 2009; Tournadre, 2014). The potential negative impacts of human generated noise on marine mammals have become a major conservation concern because of direct and suspected links between noise exposure and observed harm to marine mammals (Hildebrand, 2005). For example, naval sonar activities as well as seismic oil and gas exploration have been linked to cetacean stranding events, where necropsies have shown physical damage to vital organs (Jepson et al., 2003; Taylor et al., 2004; D Amico et al., 2009). Impacts are not limited to acute injury or death. Noise has also been shown to increase stress-hormone levels (Rolland et al., 2012), reduce foraging activity (Nowacek et al., 2004; International Whaling Commission, 2007), alter migration routes (Tyack and Clark, 1998), mask communication sounds (Clark et al., 2009), and displace marine mammals from primary feeding and breeding grounds (Jones et al., 1994; Weller et al., 2006; Tyack et al., 2011). Chronic and behavioral effects (i.e. changes in diving, feeding, social behavior) of sound exposure are difficult to study and therefore poorly understood. However, the potential for cumulative effects of even short duration responses may have profound implications for individual health, foraging and reproductive success, and potentially population trends. U.S. regulatory agencies, such as NOAA s National Marine Fisheries Service, need to better understand the effects of noise on marine mammals in order to implement effective management and protection under the Marine Mammal Protection Act (MMPA) and Endangered Species Act (ESA). Levels of harassment outlined by MMPA and ESA are used by regulatory agencies to determine if certain activities violate these protective laws, but because current data are limited, these risk functions outlining what sound exposure levels cause particular degrees of harassment (physical or behavioral) are potentially inadequate (Gilchrest et al., 2008). The Population Consequences of Acoustic Disturbance (PCAD) model was developed by the National
38 27 Research Council (2005) to examine current research status and identify research priorities for better understanding the effects of anthropogenic noise on marine mammals. This model links properties of the initial sound to population level effects through four transfer functions. The link between immediate interruption or alteration of life functions to long term effects of disturbance on individual vital rates is still poorly understood and was identified as the research area least understood (National Research Council, 2005). Behavioral response studies (BRS), using a controlled exposure experiment (CEE) paradigm, have emerged as an effective tool to study discrete behavioral responses of individual, free-ranging marine mammals to particular sounds and inform regulatory agencies of the potentially negative effects of anthropogenic sound sources on the animals (Tyack, 2009; Southall et al., 2012). These studies use animal-mounted archival tags and visual observations (focal follows) to measure an individual s response to a controlled sound exposure from a nearby ship. Typically these studies involve tagging an animal with a tag containing a passive acoustic recorder and sensors that record animal movement data (tri-axial accelerometers, magnetometers, and pressure sensors), then using a ship-mounted sound source to project a high source level sound to the animal. Behavioral observations of swim speed, dive depth, dive duration, heading, vocal behavior, group spacing, ascent and descent rate, and other metrics are made before, during and after the exposure (e.g. Curé et al., 2013; DeRuiter et al., 2013; Goldbogen et al., 2013; Antunes et al., 2014; Curé et al., 2015). The archival tag is released up to 24 hours after attachment and recovered. In situations where tags are not available, visual observations are made from a small boat before, during and after the exposure. However, these valuable studies have associated limitations and risks. They are expensive and difficult to conduct. There is limited control over exposure levels because both the target animal and sound source are moving independently. Broadcasting of high intensity signals leads to potential harassment of nearby animals that are not targeted. Furthermore, animal-mounted tags that record movement data and sound are difficult to deploy on cetaceans and do not stay on for more than a day so there is limited sample size and time scale (minutes to hours). There is a clear need to
39 28 develop additional methods to study effects of noise on marine mammals (Tyack et al., 2003; Southall et al., 2009). Combining the sound source with the behavioral sensors and acoustic recorder into a single animal-mounted tag could potentially allow for an alternative BRS method that overcomes some of the limitations of the current configuration. Such a tag would allow for better control of exposure sound levels and long-term deployments, enabling investigation of the effects of varied sound levels, repeated exposure and habituation, in a more cost effective package. The northern elephant seal, Mirounga angustirostris (Gill, 1866), presents an ideal study species to test effectiveness of such a tag. Northern elephant seals are, in comparison to cetaceans, easily accessible as they haul out in known locations to breed and molt twice a year (Worthy et al., 1992; Le Boeuf, 1994). Previous studies have shown that carrying instruments as large as 5.6 kg (mass in air) or 36 cm long does not inhibit the ability of juvenile seals (1-2 years old, kg) to swim or forage (Fletcher et al., 1996; Burgess et al., 1998). Instrument recovery rate for animals one year or older is 90-95% (Le Boeuf, 1994; Fletcher et al., 1996). They are deep diving marine mammals, with mean dive depths of over 300 m for adult males and over 500 m for adult females and maximum depths of over 1500 m (adult male) and over 1700 m (adult female) (Le Boeuf, 1994; Robinson et al., 2012), similar to many of the cetacean species thought to be most affected by anthropogenic noise (i.e. beaked whales). Although they are currently not threatened or endangered, northern elephant seals are known to regularly dive as deep as or deeper than the deep sound channel, a vertically stratified section of the water column where sound is able to propagate over long distances with little attenuation due to refraction of sound waves towards the channel axis where sound speed is at its minimum (Ewing and Worzel, 1948). Additionally, northern elephant seals have the best underwater hearing sensitivity, over the widest frequency range, of all measured phocids (Kastak and Schusterman, 1999; Reichmuth et al., 2013). This makes them a species of concern with respect to anthropogenic noise (Kastak and Schusterman, 1999), which is generally in the lower frequency range and can propagate long distances through the deep sound channel.
40 29 Elephant seal diving behavior is well studied and found to be highly stereotypic, with almost continuous deep diving by all age classes (Le Boeuf et al., 1989; Le Boeuf, 1994). Juveniles older than 2 years of age exhibit dive patterns similar to adults, spending 88% of their time at sea submerged with mean dive depths in excess of 400 m, so can serve as a proxy for adults while being easier to work with due to smaller size (Le Boeuf, 1994; Le Boeuf et al., 1996). Avoidance of shallow-water predators (i.e. killer whales and white sharks) has been suggested as one reason for the continuous deep diving behavior of northern elephant seals (Le Boeuf and Crocker, 1996). A discrete predator avoidance response has not yet been measured but would likely include changes in orientation, altered swim speed, and changes in dive state such as changing from ascending to descending or diving at a steeper rate to deeper depths, which could be inferred from sensors that measure changes in pressure, acceleration, strength of magnetic field, and turn rate (Le Boeuf and Crocker, 1996). Such a behavioral response to a perceived threat has implications for energy expenditure and aerobic dive limit, as northern elephant seals are typically highly efficient divers, utilizing negative buoyancy to performed prolonged glides to depth (Davis et al., 2001; Davis and Williams, 2012). Repeated interruption of this highly efficient dive pattern could affect foraging efficiency, energy balance, and ultimately even vital rates and individual survival. Here we describe a pilot study where we developed an animal-mounted active acoustic and motion sensing tag and tested it on free ranging, juvenile northern elephant seals. The main goals of this study were to [1] test whether an animal-borne acoustic tag can elicit any sort of behavioral response and if so, [2] would responses be related to signal content. Signal stimuli were chosen to represent both threatening and nonthreatening, natural and anthropogenic sounds from an elephant seal s environment and included sperm whale clicks, common dolphin whistles, killer whale whistles, killer whale clicks, simulated mid-frequency sonar and white noise. We hypothesized that sperm whale clicks, common dolphin whistles, and white noise would be perceived as non-threatening and result in either no response or a brief startle response, while killer whale whistles and clicks, and simulated sonar (which has similar tonal and frequency characteristics to vocalizations of killer whales) would be perceived as threatening and
41 30 result in a response characterized by an increase in depth - either a dive inversion during the ascending or bottom phase of a dive, a steeper dive angle during the descending phase of the dive, or increased swim speed if limited by the sea floor. Results Novel tag development A novel tag, which combined passive acoustic and animal movement sensors with an active acoustic sound playback system, was built, calibrated, and tested on northern elephant seals (Figure 2.1). This is the first behavioral response study of its kind conducted on pinnipeds using an animal-borne sound source. Seven deployments of the prototype tag on juvenile northern elephant seals resulted in controlled exposure experiments on five individuals (Table 2.1). Tag failure, either mechanical (flooding) or electrical (tag control board reset) was the cause of the two unsuccessful CEEs. Depth and GPS data were collected using commercially available biologgers concurrently attached to study individuals. All tagged individuals returned to Año Nuevo. There was no evidence of either instrumentation or drag from the tag package affecting normal swimming behavior, as dive profiles were similar to those of other translocated seals carrying smaller instrumentation. Deployments and basic dive behavior The five successful deployments were hours in duration, and animals received 2-24 exposures (Table 2.2, Figures ). Four of five animals dove along the seafloor up to depths of around 100 m until reaching the continental shelf edge, after which they exhibited deeper, transit dives with mean depths of m over the deep water of Monterey Canyon. They returned to the shallow, bottom limited dive pattern when returning to shallow water near Año Nuevo (Figure 2.8). Dives deeper than 100 m made up % of all dives. The single male seal, G5449, had the highest percent of dives deeper than 100 m (52.4%). He was the smallest study individual, but also had the longest deployment (Figure 2.8, first panel). One individual (G5520) rarely dove deeper than 100 m (5.0% of all dives), even over deep water (Figure 2.8, second panel). There was no evidence of foraging or drift dives by any
42 study individuals and observed dive patterns followed those seen in other translocated juvenile elephant seals. 31 Controlled exposure experiment Responses varied with dive state of the individual at time of exposure. All responses followed a general pattern of diving deep. Measurement of a dive inversion, or a change in depth greater than three meters over four seconds, was used to quantify response to exposures triggered during an ascent or during the bottom phase of a deep dive. Changes were normalized by maximum depth of dive to account for differences in dive depths. Descent rates before and after exposure were used to quantify response to exposures that occurred during a descent. Flow noise was measured before and after exposure to quantify response to exposures that occurred during bottom limited dives, as flow noise can be used as an indicator of swim speed. Dive inversions were observed multiple times through all animals entire dive profiles, not associated with exposures, however, changes in depth outside of exposure events were significantly smaller than those following exposure events (Table 2.3). Dive inversions following exposures during the ascending portion of a dive occurred where individuals inverted their ascent, returning to a descending posture and descended an additional m (SD ± 51.6 m) on average (Figure 2.9). Seven of nine exposures resulted in inversions where the seal dove as deep as or deeper than the deepest depth recorded for that dive prior to the exposure, and eight of nine exposures resulted in inversions greater than two standard deviations from the mean of all nonexposure inversions (Figure 2.9, Table 2.3). This dive inversion response was observed for white noise, sperm whale clicks, killer whale whistles, and simulated mid-frequency sonar exposures. This large dive inversion was not observed for the exposure to common dolphins, received by seal G6651 (Figure 2.9, G6651 Exposure 4). Change in depth, normalized by maximum depth of dive, was significantly different than change in depth not associated with exposures for G6651, the only seal which received multiple exposures during the ascent phase (Figure 2.10, Table 2.3; Anderson-Darling k-sample test, ADK standardized rank statistic (adjusted for ties) = 10.85, p<0.001, n=7 exposure inversions, seal G6651 and n=283 non-exposure inversions, seal G5449). No evidence
43 32 of habituation was observed for seal G6651 as dive inversion responses were as pronounced or more pronounced later in the deployment compared to earlier (Figure 2.11). Change in depth following single exposures to seals G5449 and G6110 suggested response to exposure. Both seals exhibited a change in depth greater than two standard deviations from the mean of all dive inversions, but these values could not be tested for significance because of the limited sample size of one exposure (Table 2.3). Seal G6651 dove to m following a single exposure during the bottom phase of a deep dive with a maximum depth of m, a 75% increase in maximum depth of dive (killer whale whistle exposure, Table 2.2). Two of three seals exhibited an increased descent rate following exposure during the descending phase of a dive deeper than 100 m (Figure 2.12; 9 of 10 total descending exposures). Seal G6651 had a greater descent rate in the 60 seconds following an exposure than the 60 seconds before an exposure, for all seven exposures received on the descent of a dive deeper than 100 m. Descent rate increased % (mean 125.2%; ± 95.8%). Seal G5449 also had a greater descent rate following both descending exposures it received (38.7% and 200% increase). Seal G6110 had a lower descent rate over one minute following exposure to sonar (72.7% decrease). For eight exposures (to three seals) that occurred in dives limited by the bathymetry, flow noise (in the 8-18 Hz band, a proxy for swim speed) increased seconds after the exposure ended (Figures 2.13 and 2.14). For two of the exposures, flow noise decreased for 0-30 seconds after exposure but by 60 seconds were above pre-exposure levels. Flow noise increased by 10.0 (± 8.0) db re 1 µpa from 0-30 seconds before exposure to after exposure (30.3% ± 14.7% increase). Hypothesized responses to threatening sounds (killer whale whistles, killer whale clicks, sonar) predicted an overall behavior of the seal escaping to deep water: inverting an ascent, descending more steeply if in a descent, or increasing swim speed. Hypothesized response to non-threating exposures (white noise, common dolphins, sperm whales) was no response. Seals exhibited the predicted response outlined in the introduction to 14 of 28 exposures. Predicted response percentages, calculated by exposure type, can be found in Table 2.4. Seals responded as predicted in 83% of killer whale whistle and sonar exposures, but only responded as predicted for one of six white
44 noise exposures and none of the five sperm whale click exposures. In total seals responded, regardless of prediction, to 23 of 28 total exposures (82%). 33 Discussion This pilot study aimed to evaluate the utility of an animal mounted sound source to conduct behavioral response studies (BRS) on free-ranging marine mammals. Field testing demonstrated the ability to unequivocally elicit and quantify behavioral responses from juvenile northern elephant seals using this new exposure technique. The proof of concept demonstrates that this tag has potential to expand current data regarding behavioral response of marine mammals to anthropogenic noise and potentially provide enhanced understanding of long-term effects, leading to better informed regulation of human generated sound in the ocean. The prototype tag performed mostly as expected, under a translocation paradigm. There was no evidence for effects of the tag on individual swimming ability or survival. Battery power was sufficient for up to nine day deployments, but only two CEEs spanned entire deployments due to issues with either the controller board or the housing. The playback control of the tag would occasionally reset itself. When this occurred, a sound was usually played out of sequence. In some cases, this reset caused the tag to stop working entirely. The authors believe this reset, and the resulting sound played, is also the most likely explanation for three non-exposure dive inversions of similar magnitude to inversions following exposures. These instances support the need for future iterations of the tag featuring continuous passive acoustic recording capabilities, which would allow for identification of both external and tag-made sounds and corresponding behavior, rather than only 15 min before and 15 min after exposure. The remaining tag failures resulted from housing integrity issues. Although the housing was rated to 1,500 m depth, changes in temperature and pressure likely caused failure of the slightly flexible Delrin pressure housing, sealed with o-rings and filled with mineral oil, allowing seawater to infiltrate into the tag and shorten the electronics. Future tags would benefit greatly from being potted in solid resin, however, the nonpermanent set up used in this trial allowed the researchers to monitor and modify tag
45 34 components, including programming of playback timing and playback tracks, as needed, during the field effort. Responses measured during CEEs were of several distinct types and were dependent on dive state at time of exposure. Most responses generally involved subjects diving deeper and/or longer following exposure than during baseline (non-exposure) periods. When an animal was ascending, it inverted its ascent and dove deep again, and when descending, the animal increased its angle of descent. If the animal was in shallow water, and limited by bathymetry, flow noise on the tag increased, an indicator for increased swim speed. These responses matched the hypothesized response of escaping predators by diving deeper (Le Boeuf and Crocker, 1996) or swimming more quickly. There was no indication of differential responses based on type of sound, however, because of limited sample size and anecdotal instances of no response, more trials are needed to conclusively rule out the ability of this tag to elicit differential responses to different stimuli. The clearest responses in this study involved dive inversions when the animal reversed an ascent and dove back to depth, most often as deep as or deeper than the initial maximum dive depth. These extended dive inversions served as strong indicators of response because, although all individuals exhibited multiple, smaller-scale dive inversions on ascents throughout a deployment, those inversions following exposures were magnitudes larger. Therefore, large inversions allowed a quantitative measure of response during ascents. The dive responses measured during ascents were similar in direction, but much greater in size, to dive variation seen naturally in another translocated juvenile northern elephant seal carrying a depth sensor (Davis et al., 2001). For seal G6651, which received multiple exposures during the ascending portion of deep dives, there were three instances where an exposure-like dive inversion was observed, with a change in depth similar to exposure inversions. Those three dive inversions could have been naturally occurring (e.g., exposure to an external alarming sound) or by an accidental exposure by the prototype tag following a reset (see tag assessment above). Continuous recording of ambient noise on future tag iterations, and longer-duration deployments, would allow for potential opportunistic measurement of
46 35 behavioral responses to naturally occurring threatening sounds such as predators, boats, or other man-made sounds. Results confirming this tag s ability to investigate specific responses to specific sound exposures were inconclusive, as pooled responses across all seals matched the predicted response only 50% of the time (Table 2.4). Perhaps more notably, seals responded to 82% of all exposures, but not to 100% of exposures, so it is possible there was some selection in response. Although an animal-mounted sound source is capable of eliciting behavioral responses in marine mammals, it is unclear if this exact instrument could be used to study natural responses to specific sounds, because of acoustic limitations of the playback apparatus (non-linear nature of the small transducers used) and strong context of having a sound source in such close proximity when the sound is heard. The exposure may have not been a natural enough sound to elicit a specific response that would occur naturally when an animal heard that sound. Reverberation of a sound over a distance changes acoustic properties of the signal received compared to the signal emitted. Seals are likely able to use this reverberation, differences in attenuation rates of high and low frequencies, and differences of arrival time at the ears to gather information about the source of a sound. So it is possible the animal could tell this sound was not realistically coming from directly behind them. Sudden onset of a loud sound may trigger a response, regardless of the sound, or how far away it is perceived as coming from. Differences in individual animals could also contribute to different responses, but in this small sample size that could not be determined. No single seal received multiple exposures to all exposure types, due to tag failures, so it is difficult to draw conclusions on all exposure types. However, each seal can be considered individually. While this must be considered somewhat anecdotally, some differential responses were observed. Seal G6651 was the only seal to receive an exposure of common dolphin whistles while ascending from depth and this was the only exposure where there was not an extreme dive inversion response measured. Seal G6651 did receive a second exposure to common dolphins while ascending, and no large dive inversion was measured, but this exposure occurred in only 16 m of water, so was excluded from analysis because of concern that the signal to noise ratio would be too low at this
47 36 shallow depth and the animal could not hear the exposure. Seal G5449 also did not respond to a common dolphin exposure, while descending, but did respond to sonar, killer whale whistles and sperm whales. It is possible the prediction that sperm whale clicks would be perceived as non-threatening was incorrect, as the two seals that received sperm whale exposures (G5449 and G6651) responded to all five of the exposures. This could be because the clicking sound is similar to killer whale clicks as an impulsive, transient sound. Seal G5520 responded to both exposures it received, of killer whale clicks and white noise, however it is thought that white noise did not actually serve as a good non-threatening exposure as it was likely a very unnatural sound. G6110 received a sonar exposure while descending, and a response was not detected using the criteria used to measure an increase in descent rate following exposure. However, looking at the dive profile, the animal actually decreases its descent rate following the exposure, then continues down at the same rate, before returning to the surface over a dive duration longer than most dives to that depth. This matches the response of Cuvier s beaked whales, which stopped fluking at the onset of exposure, which is hypothesized to be while they listened for the sound and assessed how to respond (DeRuiter et al., 2013). These examples provide evidence for the possibility of differential responses, however, due to the small sample size these nonpredicated responses could not be compared across multiple exposures of each type or multiple seals. Further, to fully understand the potential of this tag for behavioral response studies, experiments under the current configuration (external sound source) would need to be conducted on pinnipeds. Prior to this study, no such BRS had been done. Results here will enable development of specific questions and enhancements of the current methods and tools, and then directly connecting the responses seen with this tag to responses to an external sound source would link the two methods and allow a full comparison. Overall, responses elicited with this tag were similar to those found by other BRS on deep diving marine mammals. A pattern of interruption of normal dive behavior and either going or remaining deep longer was seen for two species of beaked whale exposed to simulated mid frequency sonar from an external, ship-mounted sound
48 37 source (DeRuiter et al., 2013; Stimpert et al., 2014). A third species of beaked whale responded to exposures of both sonar and killer whale sounds by interrupting foraging and performing an unusually long and shallow ascent (Tyack et al., 2011). Quantitative measures of response were more difficult for exposures that occurred during descents or during bottom limited dives. Although we found results consistent with the hypothesized response and responses observed in other deep diving marine mammals, natural variation in swim speed and descent angle limited ability to define statistical significance for those changes. Conclusions from the results provided by the prototype tag were limited due to small sample size, but even the limited results were promising for future use of this tag in studies other than BRS. This tag has potential for application in subsequent studies on elusive marine mammals, such as physiological studies of dive limits and studies of hearing ranges. The tag could, for example, be used as a mobile, programmable sound source to study frequency dependent hearing in marine mammal species that cannot be studied using captive psycho-physical methods. Little is known about hearing in many marine mammals because laboratory studies of hearing are not feasible due to constraints with keeping very large, migratory, and social species in captivity, but using controlled, incrementally increasing exposure levels of tonal sounds that trigger drastic changes in behavior could investigate differences in hearing sensitivities in previously inaccessible species. Further, this tag could be used to study physiological effects of unplanned and repeated extended deep dives (e.g. oxidative stress) which is another important research need recommended by experts in the field and regulatory agencies (National Research Council, 2005). Over short term deployments (less than 3 days), there was no sign of habituation, so the tag could be used to study repeated dives and possibly investigate duration of exposure until habituation. Combining implanted blood oxygen sensors with use of this tag would allow in situ measurements of diving responses. For species that are specialized for oxygen efficiency during prolonged, deep dives, physiological responses to changes in planned dive depth or duration could help physiologists better understand these adaptations (Davis and Williams, 2012). To support the potential use of this tag to study physiological effects of extended dives
49 38 possibly related to responses to anthropogenic noise, it is promising that the responses were similar to other deep diving species of high interest. Motion sensor data collected in this study could potentially be investigated further to better understand seal responses to specific sounds, but these data were insufficient for such analyses. As this was a pilot study, there were no clear or measureable predictions for a behavioral response in a juvenile northern elephant seal. Additional analyses of motion sensor data from this study could be used to inform additional hypotheses and future research opportunities with this instrument. Materials and methods A three-part study design was implemented, including tag development, pilot field efforts, and behavioral data analysis. Prototype tag development was completed in the winter and spring of Field efforts were conducted in collaboration with the Costa Lab at University of California, Santa Cruz and were completed in March and April Behavioral analysis was completed during 2013 and Tag development The prototype tag contained three components: an active acoustic playback system, a passive acoustic recording system, and a motion monitoring system (Figure 2.1). An OpenTag Single Board Controller (SBC, Loggerhead Instruments, Inc., Sarasota, FL, USA) controlled playbacks and contained a 3-axis accelerometer, 3-axis magnetometer, and 3-axis gyroscope. It was programmed (Loggerhead Instruments, Arduino programming language) to execute a playback sequence at a pre-determined time interval, using sound files stored on a SOMO playback module (4D Systems, Minchinbury, NSW, Australia), through an amplifier board (LM48580 Evaluation Board, Texas Instruments, Dallas, TX, USA) and two or three piezo electric ceramic cylinder transducers (Steiner & Martin, Inc, Doral, FL, USA; 26 mm external diameter x 22 internal diameter x 13 mm high, resonant frequency 43 khz ± 1.5 khz). The playback system was powered by a 3.7 V, 3,000 mah lithium-ion polymer rechargeable battery (Tenergy, Fremont, CA, USA). Playback of acoustic stimuli were programmed to trigger one at a time, according to a preset order, unless the tag reset itself, at which
50 39 point it started back at the beginning of the playback sequence. The cause of such resetting of the SBC was not resolved, but may have been caused by the formation of a ground loop due to differential ground potentials of the independent instruments linked together to build the single tagging system. Occurrence of resets was only documented when it coincided with the 30 minute duration of a programmed passive acoustic recording. Over all deployments, a reset was recorded in one instance (seal G6009), where two playbacks occurred within 22 minutes of one another. Stimuli were programmed to play every six hours for the first two seals, and every three hours for the remaining seals to increase likelihood of a subsurface exposure (Table 2.2). Six playback signals were used, and were chosen to include both natural and man-made sounds that were predicted to be either threatening or non-threatening. Playback signals included white noise, common dolphin whistles, and sperm whale clicks as a non-threatening controls and killer whale whistles, killer whale clicks, and simulated mid-frequency sonar as potentially threatening exposures (Figure 2.15). White noise and the simulated mid-frequency sonar were created using Adobe Audition 3 (Adobe System Incorporated, San Jose, CA, USA). The sperm whale clicks and common dolphin whistles were obtained and modified (to be 30 seconds in total length) from the Discovery of Sound in the Sea sound library 1. The killer whale tracks were obtained from the Vancouver Aquarium 2 and also modified to be 30 seconds in duration. An inverse function of the transducer transmitting sensitivity was generated and used to normalize each playbacks output over the entire frequency bandwidth. Original files were either.mp3 or.wav, and were converted.ad4 file format using the SOMO USB Recorder tool to be compatible with the SOMO playback module. Exposures were 30 seconds in duration to minimize the possibility the individual would realize the sound originated from the playback device on their back. Playback track source levels were measured using a calibrated hydrophone system (G.R.A.S. 42AC High Pressure Pistonphone, SPL 134 db re 20 µpa, 250 Hz, G.R.A.S. Sound & Vibration A/S, Holte, Denmark) and ranged from 121 to 132 db re 1 µpa at 1 m, depending on exposure type (Table 2.5)
51 40 A DSG acoustic recorder (Loggerhead Instruments, Inc., Sarasota, FL, USA) connected to an HTI-96-MIN hydrophone (High Tech, Inc., Long Beach, MS, USA; sensitivity db re 1 V/µPa; frequency response 2 Hz to 30 khz) was programmed to record continuously for 30 minutes starting 15 minutes prior to each scheduled playback in order to provide confirmation a playback occurred and to ensure there were no other environmental noise events occurring at the time of the playback. Recordings were sampled at 32 khz with 20 db of gain. The passive acoustic system was powered by a second 3.7v, 3,000 mah lithium-ion polymer battery (Tenergy, Fremont, CA, USA). All tag components were placed in an oil-filled Delrin housing, rated to 1,500 m depth. Clear, food grade mineral oil was used as a non-compressible and nonconductive filler so no air remained in the housing. Mineral oil density (about 0.8 g/cm 3 ) is closer to seawater than air, so it also provided better acoustic coupling from the transducers to the surrounding sea water, limiting acoustic impedance at that barrier. The tag package weighed approximately 1.5 kg (less than 1.2% total body mass of smallest seal) and was slightly negatively buoyant. The original tag design was modified following the first three deployments because the changes in pressure and temperature due to animal dives altered the density of the mineral oil in the housing. The slight flexibility of Delrin caused a pumping action to develop that moved sea water past the o-ring seals into the oil filled tag body. Flexible tubing with small air pockets was attached to the outside of the housing to allow changes in density of the mineral oil between the surface and at depth. This improved survivability of the tag. Field efforts Field translocation experiments were conducted in March and April 2012 on juvenile northern elephant seals at Año Nuevo State Park near Santa Cruz, CA. Seals were instrumented with the active acoustic tag placed on the animal s back (30-60 cm to the ears) and an ARGOS time-depth recorder (TDR) on the head (Mark10, Wildlife Computers, Redmond, WA, USA). Tags were attached by gluing a mesh base and stainless steel cable ties to the animal s fur using quick-cure adhesive (Devcon Minute Epoxy, ITW Polymers Adhesives North America, Danvers, MA, USA) and
52 41 then attaching the tag to the mesh base with metal cable ties. Depth was sampled every one second for seal G5449 and every four seconds for all other seals. Resolution of TDR s were 0.5 m and accuracy for these digital tags is rated at ± 1 m. The accelerometer, magnetometer, and gyroscope were all programmed to sample at 50 Hz. Animals were transported approximately 100 km south of Año Nuevo to Monterey, CA, and released. Animals swam back to Año Nuevo (Figure 2.2) and received sound exposures from the tag at the preprogrammed interval of three or six hours, and nonexposure dives served as control dives (Figure 2.8). Tags were recovered when seals returned to the beach. The cable ties were cut allowing removal of the tag, and the mesh and glue fell off in the following weeks during the annual molt. Behavioral analysis Passive recordings were screened for successful exposures and timing of exposures across multiple tag systems was synced. Additionally, the programmed sampling rate of 50 Hz for the OpenTag board was not the actual sampling rate achieved during deployments. Time accumulated when each individual sample was logged, resulting in up to 125 seconds of data writing time for every ten minute OpenTag file. An adjusted sampling rate was calculated by distributing the write time across each ten minute file. Using the adjusted sampling rate of (± 0.038) Hz, data could be plotted with time correctly distributed across the length of each file to reflect actual behavior over time. Depth data from the TDRs were viewed in Instrument Helper (Wildlife Computers, Redmond, WA, USA) and exported into a format to be used in MATLAB (MathWorks, Inc., Natick, MA, USA). Times were converted to ordinal date and time (MATLAB date/time) so data could be plotted across tagging systems. Dive data were corrected to correct for drift in the zero offset of the sensor using the IKNOS MATLAB toolbox (Figure 2.16; Yann Tremblay, unpublished). Drift can occur over the length of a deployment due to changes in barometric pressure and temperature, resulting in animal depth readings above the water surface. An example of a corrected and an uncorrected depth profile can be seen in Figure Dives were defined as any instance where the individual dove deeper than 4 m. This threshold was chosen to be
53 42 greater than the sum of the accuracy of the TDR (± 1 m) and a single body length of an individual. Depth profiles were used to examine possible seal responses to exposures. It was hypothesized that responses would look differently depending on the dive type or dive state, so analysis methods were divided by both maximum depth of dive and dive state. Dive shapes were compared to dive types described previously to identify potential presence of different dive types (e.g. transit vs. foraging; Le Boeuf et al., 1993). Dives were subset into deep (greater than 100 m) and shallow (100 m or less) dives. Dives above 100 m most often occurred where bathymetry was 100 m or less, and were likely limited by the seafloor, while deeper dives were likely pelagic dives, over Monterey Canyon, and not bottom limited, which may have an effect on responses measured (Figure 2.8; Tremblay and Cherel, 2000). Exposures were subset by dive phase (descent, bottom phase, ascent, and surface) using a custom made script for this data set, as traditional dive analysis tools would miss the potential responses expected. The beginning of an ascent was defined as the point at which an individual ascended for more than 4 meters over 20 seconds (an ascent rate of 0.2 m/s). Ascent starts were first marked using an automated script, then results were examined and adjusted manually to account for false positives or missed ascents (less than 10 per seal). Descents were defined as the segment from 0 m to the point at which the seal descended at a rate less than 0.1 m/s over 20 seconds. Bottom phases were defined as the segment between the descending and ascending segments. Ascent and descent rates are not measures of speed, but rather only the change in depth over change in time. Speed cannot be exactly measured in this experiment because only vertical displacement is measured by the TDR, not horizontal displacement. Exposures that occurred at depths shallower than 20 m were excluded from all analyses because ambient noise increases near the water s surface and the authors were not confident the animal could hear the low-level exposures near or at the surface, particularly when their head may have been above water to breathe while the tag remained underwater on the back. This criterion removed seal G6009 from all analyses, as both exposures were received at less than 10 m depth.
54 43 Dive inversions (Figure 2.9), were measured to determine potential responses during an ascending dive phase of a deep dive. Dive inversions were defined as a reversal from ascending to descending greater than 3 m over 4 seconds. This threshold was determined by the inflection point in a log survivor plot of changes in depth versus number of instances, which divided inversions possibly attributed to instrument noise from behavioral inversions. All inversion events for all ascending dive segments from deeper than 100 m were identified, and the change in depth and change in time from the inversion to the next maximum depth were measured. Change in depth or time at each inversion was normalized by dividing it by the maximum depth and total duration of the dive. Normalized change in depth was plotted against normalized change in time, and values measured immediately following exposures were highlighted (Figure 2.10). An Anderson-Darling k-sample (ADK) test was performed (Trujillo-Ortiz et al., 2007) to compare normalized change in depth between non-exposure dive inversions and exposure dive inversions for seal G6651, which received a sufficient number of exposures to run statistical analyses on. The ADK test compared the distribution of the seven inversions following exposures for seal G6651 to the distribution of the 283 inversions not associated with exposures for seal G5449, providing two independent random samples (Table 2.3). Seal G5449 was chosen for comparison because it had the largest sample of non-exposure inversions. The test was also performed using G6110 as the non-inversion group (n=8), and results were also significant at the 5% level. Only a single exposure occurred during the bottom phase of a deep dive. This response was similar to those observed during ascending dive phases, so the change in depth following exposure was measured and normalized by maximum depth of dive and reported. Exposures during descending dive phases of deep dives were analyzed for changes in descent rate following exposure. Descent rate was calculated by finding the difference in depth at exposure onset to depth at 60 seconds before and 60 seconds after exposure onset, and dividing by 60 seconds. A duration of 60 seconds was selected over which to calculate descent rate because it is twice as long as the exposure playback (30 seconds), in order to differentiate sustained changes in descent rate from brief startle responses just during the playback. Descent rates were compared and a
55 44 difference between descent rates before and after was calculated by subtracting the before rate from the after-exposure rate. A positive difference indicated an increase in descent rate following exposure, a negative value indicated a decrease in descent rate, and a zero value indicated no change in descent rate (Figure 2.12). Because an increase in descent rate is considered a subtle response that likely happens regularly in nonexposure dives, statistical analysis of changes in descent rate were not performed, but comparisons were reported for discussion of trends. Analysis of dives classified as shallow dives compared changes in flow noise on the passive acoustic recorder as a proxy for changes in speed, as diving deeper or reversing an ascent may not have been possible if the animal was in a shallow area and the maximum depth of dive was bottom limited. Previous studies have found an db re 1 µpa increase at very low frequencies (8-18 Hz) correspond to a doubling of current flow, regardless of tag or hydrophone design (Finger et al., 1979; Burgess et al., 1998). Average RMS in the 8-18 Hz frequency band was measured for s and 0-30 s before exposure and 0-30 and s after the exposure ended (Adobe Audition CC, Adobe Systems Incorporated, San Jose, CA, USA). The before and after periods were subset to detect shorter term changes in swim speed, but the entire 60 second period was taken into account when assessing a response (Figures 2.13 and 2.14). To investigate differential response of all seals to different stimuli, exposures were scored as either an as predicted or not as predicted response. Predicted response depended on hypothesized threatening or non-threatening nature of the exposure type, and a minimum level of response based on percent change, which varied for each dive state (ascending, bottom phase, descending, or shallow, bottom-limited dive). Predicted response to non-threatening exposures (white noise, common dolphin whistles, and sperm whale clicks) was no response for all dive states. Predicted responses for threatening exposures (killer whale whistles, killer whale clicks, and sonar) were to move away, to deeper water. For an ascending phase or bottom phase of a deep dive, this meant inverting ascent and diving deeper (greater than ± 2 standard deviations), for the descending phase this was observed as an increased descent rate (by greater than 50%), and for bottom limited dives this was observed as a relative increase in swim speed, as measured by flow noise (by greater than 10%). Proportion
56 45 of exposures that showed the predicted response were calculated for each exposure type and for all exposures across all seals. Motion sensor data from the active tag were viewed in MATLAB using a modified version of a script from Loggerhead Instruments, Inc. Accelerometer, gyroscope, and magnetometer data were displayed with the onset of exposure marked. Euler angles, which describe rotation about an axis (i.e. roll, pitch, and yaw) were calculated from the OpenTag sensor data to better visualize orientation of the seal in 3D space. No additional analyses were performed on the motion sensor data as this data set was determined to be insufficient to provide any conclusions on the kinematics or 3D behavior of elephant seals as they respond to particular sounds. Further, the changes in depth provide enough information to answer the objectives of this study.
57 List of symbols and abbreviations BRS: Behavioral response study CEE: Controlled exposure experiment ESA: Endangered Species Act MMPA: Marine Mammal Protection Act NMFS: National Marine Fisheries Service NOAA: National Oceanic and Atmospheric Association NRC: National Research Council SBC: Single Board Controller TDR: Time depth recorder 46 Acknowledgements We thank the students and volunteers of the Costa Laboratory at the University of California, Santa Cruz for their assistance in the field, and Sharon Nieukirk for her comments on the manuscript. Author contributions SF and HK were involved in securing funding, tag development, field work, analysis, and writing. MH provided field, analysis, and writing support; DC provided field assistance and permitting; DM provided tag development and analysis support; KS provided tag development and field support; LU provided field and data analysis support; DKM helped secure funding and provided tag development support; BLS provided analysis, theory, and writing support. Competing interests The authors maintain no competing financial interests for this study. Funding This project was funded by NOAA Fisheries, Office of Science and Technology, Ocean Acoustics Program and NOAA/OSU Cooperative Institute for Marine Resources Studies.
58 47 References Antunes, R., Kvadsheim, P. H., Lam, F. P. A., Tyack, P. L., Thomas, L., Wensveen, P. J. and Miller, P. J. O. (2014). High thresholds for avoidance of sonar by free-ranging long-finned pilot whales (Globicephala melas). Mar. Pollut. Bull. 83, Burgess, W. C., Tyack, P. L., Le Boeuf, B. J. and Costa, D. P. (1998). A programmable acoustic recording tag and first results from free-ranging northern elephant seals. Deep. Res. Part II Top. Stud. Oceanogr. 45, Clark, C., Ellison, W., Southall, B., Hatch, L., Van Parijs, S., Frankel, a and Ponirakis, D. (2009). Acoustic masking in marine ecosystems: intuitions, analysis, and implication. Mar. Ecol. Prog. Ser. 395, Curé, C., Antunes, R., Alves, A. C., Visser, F., Kvadsheim, P. H. and Miller, P. J. O. (2013). Responses of male sperm whales (Physeter macrocephalus) to killer whale sounds: implications for anti-predator strategies. Sci. Rep. 3, Curé, C., Sivle, L., Visser, F., Wensveen, P., Isojunno, S., Harris, C., Kvadsheim, P., Lam, F. and Miller, P. (2015). Predator sound playbacks reveal strong avoidance responses in a fight strategist baleen whale. Mar. Ecol. Prog. Ser. 526, D Amico, A., Gisiner, R. C., Ketten, D. R., Hammock, J. a., Johnson, C., Tyack, P. L. and Mead, J. (2009). Beaked whale strandings and naval exercises. Aquat. Mamm. 35, Davis, R. W. and Williams, T. M. (2012). The marine mammal dive response is exercise modulated to maximize aerobic dive duration DeRuiter, S. L., Southall, B. L., Calambokidis, J., Zimmer, W. M. X., Sadykova, D., Falcone, E. A., Friedlaender, A. S., Joseph, J. E., Moretti, D., Schorr, G. S., et al. (2013). First direct measurements of behavioural responses by Cuvier s beaked whales to mid-frequency active sonar. Biol. Lett. 9, Ewing, M. and Worzel, J. L. (1948). Long-range sound transmission. Geol. Soc. America Mem. 27, 1 32.
59 48 Finger, R. A., Abbagnaro, L. A. and Bauer, B. B. (1979). Measurements of lowvelocity flow noise on pressure and pressure gradient hydrophones. J. Acoust. Soc. Am. 65, Fletcher, S., Le Boeuf, B. J., Costa, D. P., Tyack, P. L. and Blackwell, S. B. (1996). Onboard acoustic recording from diving northern elephant seals. J. Acoust. Soc. Am. 100, Gilchrest, Y. V, Fetherston, T. N. and Neales, B. E. (2008). Assessing potential sites for undersea training ranges: the effects of active sonars on marine mammals. NUWC-NPT Tech. Rep. 11, Goldbogen, J. A., Southall, B. L., Deruiter, S. L., Calambokidis, J., Friedlaender, A. S., Hazen, E. L., Falcone, E. A., Schorr, G. S., Douglas, A., Moretti, D., et al. (2013). Blue whales respond to simulated mid-frequency military sonar. Proc. R. Soc. B Biol. Sci. 280, Hildebrand, J. A. (2005). Impacts of Anthropogenic Sound. In Marine Mammal Research: Conservation Beyond Crisis (ed. Reynolds, III, J. E., Perrin, W. F., Reeves, R. R., Montgomery, S., and Ragen, T. J.), pp Baltimore, Maryland: The John Hopkins University Press. Hildebrand, J. (2009). Anthropogenic and natural sources of ambient noise in the ocean. Mar. Ecol. Prog. Ser. 395, International Whaling Commission (2007). Report of the Scientific Committee. Annex K. Report of the standing working group on environmental concerns. J. Cetacean Res. Manag. 9, Jepson, P. D., Arbelo, M., Deaville, R., Patterson, I. A. P., Castro, P., Baker, J. R., Degollada, E., Ross, H. M., Herráez, P., Pocknell, A. M., et al. (2003). Gas-bubble lesions in stranded cetaceans. Nature 425, Jones, M., Swartz, S. and Dahlheim, M. (1994). Census of gray whale abundance in San Ignacio Lagoon: a follow-up study in response to low whale counts recorded during an acoustic playback study. Rep. No. NTIS PB to US Mar. Mammal Comm.
60 49 Kastak, D. and Schusterman, R. J. (1999). In-air and underwater hearing sensitivity of a northern elephant seal (Mirounga angustirostris). Can. J. Zool. 77, Le Boeuf, B. J. (1994). Variation in the Diving Pattern of Northern Elephant Seals with Age, Mass, Sex, and Reproductive Condition. In Elephant Seals: Population Ecology, Behavior, and Physiology (ed. Le Boeuf, B. J. and Laws, R. M.), pp Berkeley: University of California Press. Le Boeuf, B. J. and Crocker, D. E. (1996). Diving Behavior of Elephant Seals: Implications for Predator Avoidance. In Great White Sharks: The Biology of Carcharodon carcharias (ed. Klimley, A. P. and Ainley, D. G.), pp San Diego: Academic Press. Le Boeuf, B. J., Naito, Y., Huntley, A. C. and Asaga, T. (1989). Prolonged, continuous, deep diving by northern elephant seals. Can. J. Zool. 67, Le Boeuf, B. J., Crocker, D. E., Blackwell, S. B., Morris, P. A. and Thorson, P. H. (1993). Sex differences in diving and foraging behaviour of northern elephant seals. Symp. Zool. Soc. London 66, Le Boeuf, B. J., Morris, P. A., Blackwell, S. B., Crocker, D. E. and Costa, D. P. (1996). Diving behavior of juvenile northern elephant seals. Can. J. Zool. 74, National Research Council (2005). Marine mammal populations and ocean noise: determining when noise causes biologically significant effects. Washington, D.C., D.C.: U.S. National Academy of Sciences. Nowacek, D. P., Johnson, M. P. and Tyack, P. L. (2004). North Atlantic right whales (Eubalaena glacialis) ignore ships but respond to alerting stimuli. Proc. Biol. Sci. 271, Reichmuth, C., Holt, M. M., Mulsow, J., Sills, J. M. and Southall, B. L. (2013). Comparative assessment of amphibious hearing in pinnipeds. J. Comp. Physiol. A Neuroethol. Sensory, Neural, Behav. Physiol. 199, Richardson, W. J., Greene, C. R., Malme, C. I. and Thomson, D. H. (1995). Marine Mammals and Noise. San Diego: Academic Press.
61 50 Robinson, P. W., Costa, D. P., Crocker, D. E., Gallo-Reynoso, J. P., Champagne, C. D., Fowler, M. a., Goetsch, C., Goetz, K. T., Hassrick, J. L., Hückstädt, L. a., et al. (2012). Foraging behavior and success of a mesopelagic predator in the northeast Pacific Ocean: insights from a data-rich species, the northern elephant seal. PLoS One 7, e Rolland, R. M., Parks, S. E., Hunt, K. E., Castellote, M., Corkeron, P. J., Nowacek, D. P., Wasser, S. K. and Kraus, S. D. (2012). Evidence that ship noise increases stress in right whales. Proc. R. Soc. B Biol. Sci. 279, Southall, B. L., Berkson, J., Bowen, D., Brake, R., Eckman, J., Field, J., Gisner, R. C., Gregerson, S., Lang, W., Lewandowski, J., et al. (2009). Addressing the effects of human-generated sound on marine life: an integrated research plan for US federal agencies. Interagency Task Force on Anthropogenic Sound and the Marine Environment of the Joint Subcommittee on Ocean Science and Technology. Washington, D.C. Southall, B. L., Moretti, D., Abraham, B., Calambokidis, J., DeRuiter, S. L. and Tyack, P. L. (2012). Marine mammal behavioral response studies in Southern California: Advances in technology and experimental methods. Mar. Technol. Soc. J. 46, Stimpert, A. K., Deruiter, S. L., Southall, B. L., Moretti, D. J., Falcone, E. A., Goldbogen, J. A., Friedlaender, A., Schorr, G. S. and Calambokidis, J. (2014). Acoustic and foraging behavior of a tagged Baird s beaked whale (Berardius bairdii) exposed to simulated sonar. Sci. Rep. 4, 1 8. Taylor, B., Barlow, J., Pitman, R., Ballance, L., Klinger, T., DeMaster, D., Hildebrand, J., Urban, J., Palacios, D. and Mead, J. (2004). A call for research to assess risk of acoustic impact on beaked whale populations. Int. Whal. Comm. Sci. Comm. SC/56/E36,. Tournadre, J. (2014). Anthropogenic pressure on the open ocean: The growth of ship traffic revealed by altimeter data analysis. Geophys. Res. Lett. 41, Tremblay, Y. and Cherel, Y. (2000). Benthic and pelagic dives: a new foraging behaviour in rockhopper penguins. Mar. Ecol. Prog. Ser. 204,
62 51 Trujillo-Ortiz, A., Hernandez-Walls, R., Barba-Rojo, K., Cupal-Magana, L. and Zavala-Garcia, R. C. (2007). AnDarksamtest: Anderson-Darling k-sample procedure to test the hypothesis that the populations of the drawned groups are identical. A MATLAB file. Tyack, P. L. (2009). Acoustic playback experiments to study behavioral responses of free-ranging marine animals to anthropogenic sound. Mar. Ecol. Prog. Ser. 395, Tyack, P. L. and Clark, C. W. (1998). Quick-look report: Playback of lowfrequency sound to gray whales migrating past the central California coast. Tyack, P. L., Gordon, J. and Thompson, D. (2003). Controlled exposure experiments to determine the effects of noise on marine mammals. Mar. Technol. Soc. J. 37, Tyack, P. L., Zimmer, W. M. X., Moretti, D., Southall, B. L., Claridge, D. E., Durban, J. W., Clark, C. W., D Amico, A., DiMarzio, N., Jarvis, S., et al. (2011). Beaked whales respond to simulated and actual navy sonar. PLoS One 6, e Weller, D. W., Rickards, S. H., Bradford, A. L., Burdin, A. M. and Brownell Jr, R. L. (2006). The influence of 1997 sesimic surveys on the behavior of western gray whales of Sakhalin Island, Russia. IWC Sci. Comm. SC/58/E4, 12. Worthy, G. A. J., Morris, P. A., Costa, D. P. and Le Boeuf, B. J. (1992). Moult energetics of the northern elephant seal (Mirounga angustirostris). J. Zool. Soc. London 227,

63 Figure legends 52 Figure 2.1. The prototype tag was built, calibrated, and field tested on northern elephant seals. Tag components included a DSG passive acoustic recorder and hydrophone, an active acoustic preamplifier, playback module, transducers and OpenTag single board controller with a 9-axis motion sensor (accelerometer, magnetometer, and gyroscope).
64 53 Figure 2.2. Tracks of all seal deployments. Individual points are surface locations, and the track line is the straight line connection between subsequent points. Numbered black bars are approximate locations where exposures occurred, between the surface locations before and after the time of the exposure. Bathymetry data is from NOAA s NGDC, Southern California Coastal Relief Map, 1-arc second resolution.

65 54 Figure 2.3. Seal G5449 deployment. Individual points are surface locations, and the track line is the straight line connection between subsequent points. Numbered black bars are approximate locations where exposures occurred, between the surface locations before and after the time of the exposure. Bathymetry data is from NOAA s NGDC, Southern California Coastal Relief Map, 1-arc second resolution.
66 55 Figure 2.4. Seal G5520 deployment. Individual points are surface locations, and the track line is the straight line connection between subsequent points. Numbered black bars are approximate locations where exposures occurred, between the surface locations before and after the time of the exposure. Bathymetry data is from NOAA s NGDC, Southern California Coastal Relief Map, 1-arc second resolution.
67 56 Figure 2.5. Seal G6009 deployment. Individual points are surface locations, and the track line is the straight line connection between subsequent points. Numbered black bars are approximate locations where exposures occurred, between the surface locations before and after the time of the exposure. Bathymetry data is from NOAA s NGDC, Southern California Coastal Relief Map, 1-arc second resolution.
68 57 Figure 2.6. Seal G6110 deployment. Individual points are surface locations, and the track line is the straight line connection between subsequent points. Numbered black bars are approximate locations where exposures occurred, between the surface locations before and after the time of the exposure. Bathymetry data is from NOAA s NGDC, Southern California Coastal Relief Map, 1-arc second resolution.
69 58 Figure 2.7. Seal G6651 deployment. Individual points are surface locations, and the track line is the straight line connection between subsequent points. Numbered black bars are approximate locations where exposures occurred, between the surface locations before and after the time of the exposure. Bathymetry data is from NOAA s NGDC, Southern California Coastal Relief Map, 1-arc second resolution.





70 59 Figure 2.8. Depth profiles and seafloor bathymetry for complete deployments. Seal dive profile is in blue, seafloor bathymetry is in gray and exposures are indicated by black dots. Bathymetry data is from NOAA s NGDC, Southern California Coastal Relief Map, 1-arc second resolution. Instances where a dive overlaps the seafloor are an artifact of extracting bathymetry data from known GPS surface positions, which may not be available for every surfacing.
71 60 Figure 2.9. Dive profiles for all exposures received during ascending phases of dives. Black symbol indicates exposure time and type: ( ) sperm whale clicks, ( ) white noise, ( ) killer whale whistles ( ) common dolphin whistles, ( ) simulated midfrequency sonar, and (+) killer whale clicks.
72 61 Figure Plot of normalized change in depth versus normalized change in time for all dive inversions during ascending phase of all deep dives (> 100m) for seals G5449, G6110, and G6551. Dive inversions not corresponding to exposures are symbolized by dots, exposure dive inversions are symbolized based on exposure type : ( ) sperm whale clicks, ( ) white noise, ( ) killer whale whistles ( ) common dolphin whistles, ( ) simulated mid-frequency sonar, and (+) killer whale clicks..
73 62 Figure Plot of normalized change in depth versus elapsed time (in hours) for all dive inversions during the ascending phase of all deep dives (> 100m) for seal G6651. Dive inversions not corresponding to exposures are symbolized by dots, exposure dive inversions are symbolized based on exposure type: ( ) sperm whale clicks, ( ) white noise, ( ) killer whale whistles ( ) common dolphin whistles, ( ) simulated midfrequency sonar, and (+) killer whale clicks.

74 63 Figure Mean descent rates, in m/s, for seals G5449, G6110 and G seconds before and 60 after exposure. Blue bars are descent rate before exposure, red bars are descent rate over the 60 seconds after exposure. Exposure number is given for each seal and corresponds to the following exposure types: G5449- (4) killer whale whistles, (5) common dolphin whistles; G6110- (2) sonar; G6651 (2) white noise, (6) white noise, (8) common dolphin whistles, (11) white noise, (13) sperm whale clicks, (19) sonar, and (20) white noise.

75 64 Figure Spectrogram of passive acoustic recorder showing noise before, during, and after a sonar exposure (exposure 3 to seal G5449). The four broadband dark lines are simulated sonar ping exposures. Energy in the lower frequencies (< 800 Hz) is generated by flow noise, which can serve as a proxy for swim speed, where increased flow noise indicates an increase in flow noise.
76 65 Figure Noise level (relative amplitude) in the 8-18 Hz band before and after exposure, as a proxy for relative change in speed. An db re 1 µpa increase in flow noise is approximately equal to a doubling of speed (Burgess et al., 1998; Finger et al., 1979).

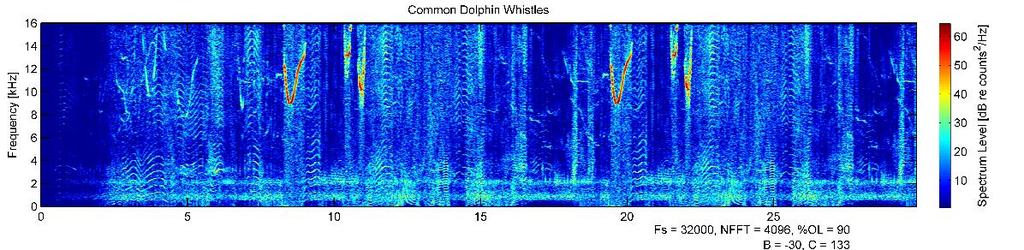
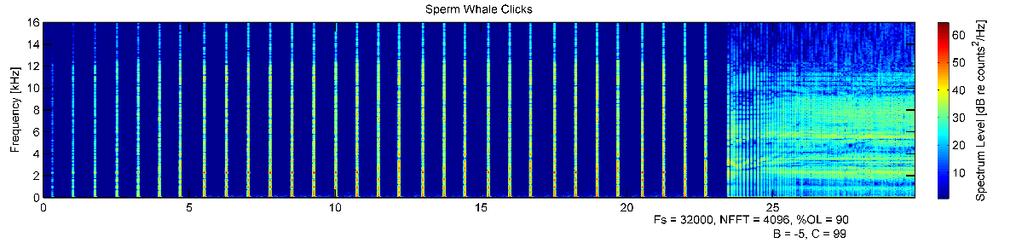
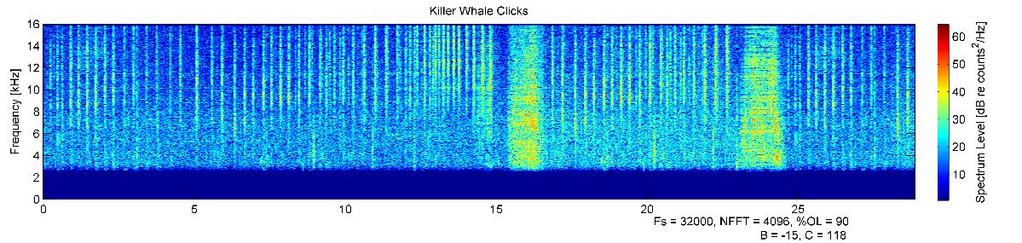
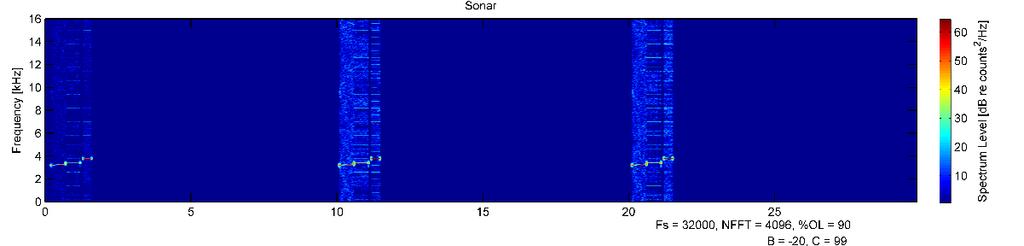
77 Figure Spectrograms of playback tracks. 66
78 67 Figure Example of depth profiles corrected for zero-offset (blue), and uncorrected (gray) for seal G6651.
 in the field by experts in elephant seal physiology, based on standard length and weight.")
79 Table 2.1. Study individual information, by seal. Age was either known from initial tagging as a pup or estimated(*) in the field by experts in elephant seal physiology, based on standard length and weight. Body mass was measured before instrumentation and translocation. Number of dives recorded is all instances where the animal dove deeper than 4 m. Mean (and standard deviation) dive duration, in minutes, is given for all dives and the subset of dives deeper than 100 m. Seal Age Sex Body Mass (kg) Deployment Duration (hours) Number of dives recorded G5449 1* M Number of dives deeper than 100m (% of total) 509 (52.4%) G F (5.0%) G F (23.9%) G6110 2* F (18.9%) G6651 1* F (35.8%) Mean dive depth (meters) All dives (± ) (± ) (± ) (± ) (± ) Dives deeper than 100 m (± ) (± ) (± ) (±165.32) (± ) Mean dive duration (seconds) All dives (± ) (± ) (± ) (± ) (± ) Dives deeper than 100 m (± ) (± ) (± ) (± ) (± ) Median dive duration (seconds) All dives Dives deeper than 100 m Tables 68
~ A Behavioral Response Study in 2007 &2008 (BRS 07/08) was conducted in the Bahamas to
 was conducted in the Bahamas to") Biological and Behavioral Response Studies in the Bahamas in 27 28 (BRS 7/8) ~ Project Summary ~ ~ A Behavioral Response Study in 27 &28 (BRS 7/8) was conducted in the Bahamas to study diving behavior
Biological and Behavioral Response Studies in the Bahamas in 27 28 (BRS 7/8) ~ Project Summary ~ ~ A Behavioral Response Study in 27 &28 (BRS 7/8) was conducted in the Bahamas to study diving behavior
Biological & Behavioral Response Studies of. Marine Mammals in Southern California, BRS.org. Photos taken under NMFS permit #
 Biological & Behavioral Response Studies of Marine Mammals in Southern California, 2015 ( SOCAL- 15) www.socal- BRS.org Photos taken under NMFS permit #14534-2 1 SOCAL- 15 OVERVIEW SOCAL- 15 continues
Biological & Behavioral Response Studies of Marine Mammals in Southern California, 2015 ( SOCAL- 15) www.socal- BRS.org Photos taken under NMFS permit #14534-2 1 SOCAL- 15 OVERVIEW SOCAL- 15 continues
Marine Mammals. James M. Price. Division of Environmental Sciences. from NOAA photograph library
 Marine Mammals from NOAA photograph library James M. Price James.Price@boem.gov Division of Environmental Sciences Environmental Studies Program R/V Pisces (from NOAA Web site) Develops and oversees applied
Marine Mammals from NOAA photograph library James M. Price James.Price@boem.gov Division of Environmental Sciences Environmental Studies Program R/V Pisces (from NOAA Web site) Develops and oversees applied
Fine-scale Focal DTAG Behavioral Study in the Gulf of Maine
 DISTRIBUTION STATEMENT A: Approved for public release; distribution is unlimited. Fine-scale Focal DTAG Behavioral Study in the Gulf of Maine Alessandro Bocconcelli Woods Hole Oceanographic Institution
DISTRIBUTION STATEMENT A: Approved for public release; distribution is unlimited. Fine-scale Focal DTAG Behavioral Study in the Gulf of Maine Alessandro Bocconcelli Woods Hole Oceanographic Institution
Marine Mammal Acoustic Tracking from Adapting HARP Technologies
 DISTRIBUTION STATEMENT A: Approved for public release; distribution is unlimited. Marine Mammal Acoustic Tracking from Adapting HARP Technologies Sean M. Wiggins and John A. Hildebrand Marine Physical
DISTRIBUTION STATEMENT A: Approved for public release; distribution is unlimited. Marine Mammal Acoustic Tracking from Adapting HARP Technologies Sean M. Wiggins and John A. Hildebrand Marine Physical
Fine-scale Focal Dtag Behavioral Study of Diel Trends in Activity Budgets and Sound Production of Endangered Baleen Whales in the Gulf of Maine
 Fine-scale Focal Dtag Behavioral Study of Diel Trends in Activity Budgets and Sound Production of Endangered Baleen Whales in the Gulf of Maine Susan E. Parks The Pennsylvania State University, Applied
Fine-scale Focal Dtag Behavioral Study of Diel Trends in Activity Budgets and Sound Production of Endangered Baleen Whales in the Gulf of Maine Susan E. Parks The Pennsylvania State University, Applied
Using DTAGs to understand sound use, behavior, and vessel and associated noise effects in Southern Resident killer whales
 Western Washington University Western CEDAR Salish Sea Ecosystem Conference 2018 Salish Sea Ecosystem Conference (Seattle, Wash.) Apr 6th, 9:15 AM - 9:30 AM Using DTAGs to understand sound use, behavior,
Western Washington University Western CEDAR Salish Sea Ecosystem Conference 2018 Salish Sea Ecosystem Conference (Seattle, Wash.) Apr 6th, 9:15 AM - 9:30 AM Using DTAGs to understand sound use, behavior,
Final Report Alaska Department of Fish and Game State Wildlife Grant T July 1, 2003 June 30, 2006:
 Final Report Alaska Department of Fish and Game State Wildlife Grant T-1-6-4 July 1, 2003 June 30, 2006: Acoustic Monitoring for Killer Whales in the Bering Sea John Hildebrand and Erin Oleson Marine Physical
Final Report Alaska Department of Fish and Game State Wildlife Grant T-1-6-4 July 1, 2003 June 30, 2006: Acoustic Monitoring for Killer Whales in the Bering Sea John Hildebrand and Erin Oleson Marine Physical
Lifeforce Foundation Orca Conservation Programs
 Lifeforce Foundation Orca Conservation Programs Photo Captions: Start Top left Clockwise 1. Over fishing and entanglement in fishing nets and other debris is a threat to orcas. 2. Boaters should be aware
Lifeforce Foundation Orca Conservation Programs Photo Captions: Start Top left Clockwise 1. Over fishing and entanglement in fishing nets and other debris is a threat to orcas. 2. Boaters should be aware
Sound production in mysticetes. Background. Cetacean vocal frequency ranges Dolphins
 Sound production in mysticetes Fish 475 - May 4 2007 Background Cetaceans produce sound in the widest range of frequencies for any Class of animal Subsonic to supersonic and everywhere in between Sound
Sound production in mysticetes Fish 475 - May 4 2007 Background Cetaceans produce sound in the widest range of frequencies for any Class of animal Subsonic to supersonic and everywhere in between Sound
Jeopardy. Diving Acoustics Survey Misc. Q $100 Q $100 Q $100 Q $100 Q $200 Q $200 Q $200 Q $200 Q $300 Q $300 Q $300 Q $300
 Jeopardy ID, Skins, & Skulls Q $100 Q $200 Q $300 Q $400 Q $500 Diving Acoustics Survey Misc. Q $100 Q $100 Q $100 Q $100 Q $200 Q $200 Q $200 Q $200 Q $300 Q $300 Q $300 Q $300 Q $400 Q $400 Q $400 Q
Jeopardy ID, Skins, & Skulls Q $100 Q $200 Q $300 Q $400 Q $500 Diving Acoustics Survey Misc. Q $100 Q $100 Q $100 Q $100 Q $200 Q $200 Q $200 Q $200 Q $300 Q $300 Q $300 Q $300 Q $400 Q $400 Q $400 Q
I. Monitoring and Adaptive Management Approach. Potential Effects, Monitoring Studies & Mitigation
 I. Monitoring and Adaptive Management Approach Define the problem or management objective. Determine potential effects of the project on indicator species. Emphasis on examining potentially negative, rather
I. Monitoring and Adaptive Management Approach Define the problem or management objective. Determine potential effects of the project on indicator species. Emphasis on examining potentially negative, rather
Assessment of cumulative Sound Exposure Levels (SEL) for marine piling events
 for marine piling events") Assessment of cumulative Sound Exposure Levels (SEL) for marine piling events P A Lepper 1, S P Robinson 2, M A Ainslie 3, P D Theobald 2, C A F de Jong 4 1 Loughborough University, Leicestershire, U K,
Assessment of cumulative Sound Exposure Levels (SEL) for marine piling events P A Lepper 1, S P Robinson 2, M A Ainslie 3, P D Theobald 2, C A F de Jong 4 1 Loughborough University, Leicestershire, U K,
Behavioral and Physiological Response of Baleen Whales to Ships and Ship Noise
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Behavioral and Physiological Response of Baleen Whales to Ships and Ship Noise John Calambokidis Cascadia Research Collective
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Behavioral and Physiological Response of Baleen Whales to Ships and Ship Noise John Calambokidis Cascadia Research Collective
Evidence of Anton van Helden in the matter of the applications by Trans Tasman Resources Limited for marine and discharge consents to recover iron
 Evidence of Anton van Helden in the matter of the applications by Trans Tasman Resources Limited for marine and discharge consents to recover iron sand in the South Taranaki Bight Qualifications of Anton
Evidence of Anton van Helden in the matter of the applications by Trans Tasman Resources Limited for marine and discharge consents to recover iron sand in the South Taranaki Bight Qualifications of Anton
CLIMATE VARIABILITY IS LINKED TO DIET SWITCHING IN A
 CLIMATE VARIABILITY IS LINKED TO DIET SWITCHING IN A MARINE PREDATOR, THE NORTHERN ELEPHANT SEAL Chandra Goetsch 1, Melinda Conners 1, Yoko Mitani 2, William Walker 3, Samantha Simmons 4, Colleen Reichmuth
CLIMATE VARIABILITY IS LINKED TO DIET SWITCHING IN A MARINE PREDATOR, THE NORTHERN ELEPHANT SEAL Chandra Goetsch 1, Melinda Conners 1, Yoko Mitani 2, William Walker 3, Samantha Simmons 4, Colleen Reichmuth
2nd Technical Research Paper Converted to Docent Friendly Summary Status. October 9, 2013
 2nd Technical Research Paper Converted to Docent Friendly Summary Status October 9, 2013 The following is a docent friendly summary of a Northern Elephant Seal (Mirounga augustirostis) Research Paper:
2nd Technical Research Paper Converted to Docent Friendly Summary Status October 9, 2013 The following is a docent friendly summary of a Northern Elephant Seal (Mirounga augustirostis) Research Paper:
Behavioral and physiological response of baleen whales to ships and ship noise
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Behavioral and physiological response of baleen whales to ships and ship noise John Calambokidis Cascadia Research Collective
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Behavioral and physiological response of baleen whales to ships and ship noise John Calambokidis Cascadia Research Collective
National Report on Large Whale Entanglements
 National Report on Large Whale Entanglements Confirmed in the United States in 2017 In 2017, 76 confirmed cases of large whale entanglements were documented along the coasts of the United States. Seventy
National Report on Large Whale Entanglements Confirmed in the United States in 2017 In 2017, 76 confirmed cases of large whale entanglements were documented along the coasts of the United States. Seventy
2016 West Coast Entanglement Summary
 March 2017 2016 West Coast Entanglement Summary Overview of Entanglement Data west coast region MMHSRP Permit #18786 In 2016, 71 separate cases of entangled whales were reported off the coasts of Washington,
March 2017 2016 West Coast Entanglement Summary Overview of Entanglement Data west coast region MMHSRP Permit #18786 In 2016, 71 separate cases of entangled whales were reported off the coasts of Washington,
Oregon Hatchery Research Center January 2014 David L. G. Noakes, Professor & Director
 Oregon Hatchery Research Center January 2014 David L. G. Noakes, Professor & Director Research Proposal Homing Homing behavior is a striking feature of the biology of Pacific salmon, and is the basis for
Oregon Hatchery Research Center January 2014 David L. G. Noakes, Professor & Director Research Proposal Homing Homing behavior is a striking feature of the biology of Pacific salmon, and is the basis for
Tagging and Playback Studies to Toothed Whales
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited Tagging and Playback Studies to Toothed Whales Peter L. Tyack Biology Department Woods Hole Oceanographic Institution Woods
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited Tagging and Playback Studies to Toothed Whales Peter L. Tyack Biology Department Woods Hole Oceanographic Institution Woods
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited.
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Examination of Health Effects and Long-Term Impacts of Deployments of Multiple Tag Types on Blue, Humpback, and Gray Whales
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Examination of Health Effects and Long-Term Impacts of Deployments of Multiple Tag Types on Blue, Humpback, and Gray Whales
BEFORE THE ENVIRONMENTAL PROTECTION AUTHORITY JOINT STATEMENT OF EXPERTS IN THE FIELD OF MARINE MAMMALS
 BEFORE THE ENVIRONMENTAL PROTECTION AUTHORITY IN THE MATTER of the Exclusive Economic Zone and Continental Shelf (Environmental Effects) Act 2012 AND IN THE MATTER of an Application for Marine D u m p
BEFORE THE ENVIRONMENTAL PROTECTION AUTHORITY IN THE MATTER of the Exclusive Economic Zone and Continental Shelf (Environmental Effects) Act 2012 AND IN THE MATTER of an Application for Marine D u m p
Cetacean Identification Key
 Cetacean Identification Key KAI Marine Services LIFE+ Migrate Short beaked common dolphin (Delphinus delphis) Pale yellow and grey hourglass pattern on the sides Black back Grey rear flanks Frequent jumps
Cetacean Identification Key KAI Marine Services LIFE+ Migrate Short beaked common dolphin (Delphinus delphis) Pale yellow and grey hourglass pattern on the sides Black back Grey rear flanks Frequent jumps
Pacific Missile Range Facility (PMRF) Species Verification and Satellite Tagging Test: January Post-Test Report
 Species Verification and Satellite Tagging Test: January Post-Test Report") Pacific Missile Range Facility (PMRF) Species Verification and Satellite Tagging Test: January 2012 Post-Test Report POC: David J. Moretti Naval Undersea Warfare Center Division Newport: Marine Mammal
Pacific Missile Range Facility (PMRF) Species Verification and Satellite Tagging Test: January 2012 Post-Test Report POC: David J. Moretti Naval Undersea Warfare Center Division Newport: Marine Mammal
Acoustic Seaglider for Beaked Whale Detection
 DISTRIBUTION STATEMENT A: Approved for public release; distribution is unlimited. Acoustic Seaglider for Beaked Whale ion Neil M. Bogue and Jim Luby Applied Physics Laboratory University of Washington
DISTRIBUTION STATEMENT A: Approved for public release; distribution is unlimited. Acoustic Seaglider for Beaked Whale ion Neil M. Bogue and Jim Luby Applied Physics Laboratory University of Washington
Baseline Behavior of Pilot Whales and their Responses to Playback of Anthropogenic and Natural Sounds
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Baseline Behavior of Pilot Whales and their Responses to Playback of Anthropogenic and Natural Sounds Peter Tyack, Senior
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Baseline Behavior of Pilot Whales and their Responses to Playback of Anthropogenic and Natural Sounds Peter Tyack, Senior
Human-generated sound and marine mammals
 feature Human-generated sound and marine mammals Peter L. Tyack Loud anthropogenic noises can alter the behavior of whales and other marine mammals, sometimes with fatal consequences. Peter Tyack is a
feature Human-generated sound and marine mammals Peter L. Tyack Loud anthropogenic noises can alter the behavior of whales and other marine mammals, sometimes with fatal consequences. Peter Tyack is a
Southern Resident Killer Whale Information Webex
 Southern Resident Killer Whale Information Webex February 23 rd, 2018 Sheila J Thornton, Ph.D Research Scientist, Fisheries and Oceans Canada Photo: S.J. Thornton Resident Killer Whale Biology Resident
Southern Resident Killer Whale Information Webex February 23 rd, 2018 Sheila J Thornton, Ph.D Research Scientist, Fisheries and Oceans Canada Photo: S.J. Thornton Resident Killer Whale Biology Resident
Fine Scale Baleen Whale Behavior Observed Via Tagging Over Daily Time Scales
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Fine Scale Baleen Whale Behavior Observed Via Tagging Over Daily Time Scales Mark Baumgartner Woods Hole Oceanographic
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Fine Scale Baleen Whale Behavior Observed Via Tagging Over Daily Time Scales Mark Baumgartner Woods Hole Oceanographic
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited.
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Off-Range Beaked Whale Studies (ORBS): Baseline Data and Tagging Development for Northern Bottlenose Whales (Hyperoodon
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Off-Range Beaked Whale Studies (ORBS): Baseline Data and Tagging Development for Northern Bottlenose Whales (Hyperoodon
Anthropogenic Noise and the Marine Environment
 Anthropogenic Noise and the Marine Environment R. Hillson and H.-J. Shyu Information Technology Division Introduction: The impact of anthropogenic noise on the marine environment is a subject of increasing
Anthropogenic Noise and the Marine Environment R. Hillson and H.-J. Shyu Information Technology Division Introduction: The impact of anthropogenic noise on the marine environment is a subject of increasing
Providing Field Support for the Behavior Response Study (BRS-07)
") Providing Field Support for the Behavior Response Study (BRS-07) Diane Claridge Bahamas Marine Mammal Research Organization P.O. Box AB-20714, Marsh Harbour Abaco, Bahamas phone: (242) 366-4155 fax: (242)
Providing Field Support for the Behavior Response Study (BRS-07) Diane Claridge Bahamas Marine Mammal Research Organization P.O. Box AB-20714, Marsh Harbour Abaco, Bahamas phone: (242) 366-4155 fax: (242)
Puget Sound s whales face intertwined obstacles By The Seattle Times, adapted by Newsela staff Jul. 15, :00 AM
 Puget Sound s whales face intertwined obstacles By The Seattle Times, adapted by Newsela staff Jul. 15, 2014 4:00 AM A sheet of water cascades off the back of an Orca that surfaced in Haro Strait near
Puget Sound s whales face intertwined obstacles By The Seattle Times, adapted by Newsela staff Jul. 15, 2014 4:00 AM A sheet of water cascades off the back of an Orca that surfaced in Haro Strait near
Puget Sound's whales face intertwined obstacles
 Puget Sound's whales face intertwined obstacles By The Seattle Times, adapted by Newsela staff on 07.15.14 Word Count 892 A sheet of water cascades off the back of an Orca that surfaced in Haro Strait
Puget Sound's whales face intertwined obstacles By The Seattle Times, adapted by Newsela staff on 07.15.14 Word Count 892 A sheet of water cascades off the back of an Orca that surfaced in Haro Strait
Status of humpback, blue, and gray whales along the US West Coast and relevance to entanglements
 Status of humpback, blue, and gray whales along the US West Coast and relevance to entanglements John Calambokidis Research Biologist Cascadia Research Goals of talk Updated status of humpback, blue and
Status of humpback, blue, and gray whales along the US West Coast and relevance to entanglements John Calambokidis Research Biologist Cascadia Research Goals of talk Updated status of humpback, blue and
Underwater sound in relation to dredging
 CEDA Underwater sound in relation to dredging Dr. Frank Thomsen DHI Group Underwater sound and dredging http://www.dredging.org/, special thanks to Anna Csiti 23 4 2013 2 Outline Sound and marine life
CEDA Underwater sound in relation to dredging Dr. Frank Thomsen DHI Group Underwater sound and dredging http://www.dredging.org/, special thanks to Anna Csiti 23 4 2013 2 Outline Sound and marine life
Patterns of Southern Resident Killer Whale (Orcinus orca) Movement in Relation to the Tides and Currents
 Movement in Relation to the Tides and Currents") Patterns of Southern Resident Killer Whale (Orcinus orca) Movement in Relation to the Tides and Currents Erin M. Soucy Beam Reach Marine Science and Sustainability School 620 University Road Friday Harbor,
Patterns of Southern Resident Killer Whale (Orcinus orca) Movement in Relation to the Tides and Currents Erin M. Soucy Beam Reach Marine Science and Sustainability School 620 University Road Friday Harbor,
Effect on Marine Life by Noise of Offshore Wind Farm S.JIANG 1 & J.P. HOU 1
 5th International Conference on Advanced Design and Manufacturing Engineering (ICADME 2015) Effect on Marine Life by Noise of Offshore Wind Farm S.JIANG 1 & J.P. HOU 1 1 Third Institute of Oceangraphy,SOA,Xiamen,Fujian,China
5th International Conference on Advanced Design and Manufacturing Engineering (ICADME 2015) Effect on Marine Life by Noise of Offshore Wind Farm S.JIANG 1 & J.P. HOU 1 1 Third Institute of Oceangraphy,SOA,Xiamen,Fujian,China
Remote Monitoring of Dolphins and Whales in the High Naval Activity Areas in Hawaiian Waters
 DISTRIBUTION STATEMENT A: Approved for public release; distribution is unlimited. Remote Monitoring of Dolphins and Whales in the High Naval Activity Areas in Hawaiian Waters Whitlow W. L. Au and Marc
DISTRIBUTION STATEMENT A: Approved for public release; distribution is unlimited. Remote Monitoring of Dolphins and Whales in the High Naval Activity Areas in Hawaiian Waters Whitlow W. L. Au and Marc
WODA Technical Guidance on Underwater Sound in Relation to Dredging
 WODA Technical Guidance on Underwater Sound in Relation to Dredging Frank Thomsen, Fabrizio Borsani, Pim de Wit, Douglas Clarke, Christ de Jong, Frederik Goethals, Martine Holtkamp, Gerard van Raalte,
WODA Technical Guidance on Underwater Sound in Relation to Dredging Frank Thomsen, Fabrizio Borsani, Pim de Wit, Douglas Clarke, Christ de Jong, Frederik Goethals, Martine Holtkamp, Gerard van Raalte,
Baseline Behavior of Pilot Whales and their Responses to Playback of Anthropogenic and Natural Sounds
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Baseline Behavior of Pilot Whales and their Responses to Playback of Anthropogenic and Natural Sounds Peter Tyack Woods
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Baseline Behavior of Pilot Whales and their Responses to Playback of Anthropogenic and Natural Sounds Peter Tyack Woods
Use of a low power, airgun sound source to accurately determine sound. Transmission Loss characteristics at the proposed Robin Rigg. Windfarm site.
 Submitted to: Submitted by: The Scottish Executive On behalf of: Mr M J Swanwick Mr S J Parvin EON-UK Subacoustech Ltd Westwood Way Chase Mill Westwood Business Park Winchester Road Coventry Bishop s Waltham
Submitted to: Submitted by: The Scottish Executive On behalf of: Mr M J Swanwick Mr S J Parvin EON-UK Subacoustech Ltd Westwood Way Chase Mill Westwood Business Park Winchester Road Coventry Bishop s Waltham
Concept for a Whale Protection Zone for the Endangered Southern Resident Killer Whale
 Concept for a Whale Protection Zone for the Endangered Southern Resident Killer Whale Supported (in part) by the Charlotte Martin Foundation and the Norcliffe Foundation The concept described in this document
Concept for a Whale Protection Zone for the Endangered Southern Resident Killer Whale Supported (in part) by the Charlotte Martin Foundation and the Norcliffe Foundation The concept described in this document
BAJA BLUE WHALE RESEARCH 2011
 BAJA BLUE WHALE RESEARCH 2011 Summary of research conducted by Cascadia Research as a part of an Oceanic Society Research Expedition with volunteers from Road Scholar Expedition Dates: ch 26 il 2, 2011
BAJA BLUE WHALE RESEARCH 2011 Summary of research conducted by Cascadia Research as a part of an Oceanic Society Research Expedition with volunteers from Road Scholar Expedition Dates: ch 26 il 2, 2011
Evaluating the Effects of Stressors on Immune Function during Simulated Dives in Marine Mammals
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Evaluating the Effects of Stressors on Immune Function during Simulated Dives in Marine Mammals Tracy Romano and Laura
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Evaluating the Effects of Stressors on Immune Function during Simulated Dives in Marine Mammals Tracy Romano and Laura
Fine-Scale Survey of Right and Humpback Whale Prey Abundance and Distribution
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Fine-Scale Survey of Right and Humpback Whale Prey Abundance and Distribution Joseph D. Warren School of Marine and Atmospheric
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Fine-Scale Survey of Right and Humpback Whale Prey Abundance and Distribution Joseph D. Warren School of Marine and Atmospheric
Acoustic and Visual Survey of Cetaceans at Palmyra Atoll
 Acoustic and Visual Survey of Cetaceans at Palmyra Atoll Trip report 08/2010 Palmyra, August 18 September 9, 2010 Jason P Larese Mark Deakos Contact: sbaumann@ucsd.edu, jhildebrand@ucsd.edu Scripps Institution
Acoustic and Visual Survey of Cetaceans at Palmyra Atoll Trip report 08/2010 Palmyra, August 18 September 9, 2010 Jason P Larese Mark Deakos Contact: sbaumann@ucsd.edu, jhildebrand@ucsd.edu Scripps Institution
DECEMBER 15-18, 1997 ADELAIDE, SOUTH AUSTRALIA. Douglas H. Cato. Defence Science and Technology Organisation PO Box 44, Pyrmont, NSW, 2009, Australia
 FIFTH INTERNATIONAL CONGRESS ON SOUND AND VIBRATION DECEMBER 15-18, 1997 ADELAIDE, SOUTH AUSTRALIA AMBIENT SEA NOISE IN AUSTRALIAN WATERS Douglas H. Cato Defence Science and Technology Organisation PO
FIFTH INTERNATIONAL CONGRESS ON SOUND AND VIBRATION DECEMBER 15-18, 1997 ADELAIDE, SOUTH AUSTRALIA AMBIENT SEA NOISE IN AUSTRALIAN WATERS Douglas H. Cato Defence Science and Technology Organisation PO
Species and Area Protection with Regards to Offshore Wind Farms. Dr. Folchert R. van Dijken
 Species and Area Protection with Regards to Offshore Wind Farms Dr. Folchert R. van Dijken 25 January 2012 2 Economic activities on the NL Continental shelf Shipping - 260,000 movements - Seaport like
Species and Area Protection with Regards to Offshore Wind Farms Dr. Folchert R. van Dijken 25 January 2012 2 Economic activities on the NL Continental shelf Shipping - 260,000 movements - Seaport like
3S 2 : Behavioral Response Studies of Cetaceans to Navy Sonar Signals in Norwegian Waters
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. 3S 2 : Behavioral Response Studies of Cetaceans to Navy Sonar Signals in Norwegian Waters Patrick Miller Sea Mammal Research
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. 3S 2 : Behavioral Response Studies of Cetaceans to Navy Sonar Signals in Norwegian Waters Patrick Miller Sea Mammal Research
Habitat Use and Behavioral Monitoring of Hawaiian Monk Seals in Proximity to the Navy Hawaii Range Complex
 Project Annual Report Habitat Use and Behavioral Monitoring of Hawaiian Monk Seals in Proximity to the Navy Hawaii Range Complex Report Period: July 2011 June 2012 Submitted by: Kenady Wilson, Scientist
Project Annual Report Habitat Use and Behavioral Monitoring of Hawaiian Monk Seals in Proximity to the Navy Hawaii Range Complex Report Period: July 2011 June 2012 Submitted by: Kenady Wilson, Scientist
AGENCY: National Marine Fisheries Service (NMFS), National Oceanic and Atmospheric
, National Oceanic and Atmospheric") This document is scheduled to be published in the Federal Register on 05/30/2013 and available online at http://federalregister.gov/a/2013-12869, and on FDsys.gov BILLING CODE 3510-22-P DEPARTMENT OF COMMERCE
This document is scheduled to be published in the Federal Register on 05/30/2013 and available online at http://federalregister.gov/a/2013-12869, and on FDsys.gov BILLING CODE 3510-22-P DEPARTMENT OF COMMERCE
Review of new information on pollution, underwater sound and disturbance: Military, including munitions
 ASCOBANS 14 th Advisory Committee Meeting Document AC14/Doc. 27(O) San Sebastián, Spain, 19 21 April 2007 Dist. 19 April 2007 Agenda Item 6.5.2: Review of new information on pollution, underwater sound
ASCOBANS 14 th Advisory Committee Meeting Document AC14/Doc. 27(O) San Sebastián, Spain, 19 21 April 2007 Dist. 19 April 2007 Agenda Item 6.5.2: Review of new information on pollution, underwater sound
Underwater noise and offshore windfarms
 Underwater noise and offshore windfarms Dr Jeremy Nedwell, Mr John Langworthy and Mr Daren Howell BWEA Conference 4/3/04 Subacoustech reference: 544R0503, COWRIE Copyright. Aim of study To evaluate the
Underwater noise and offshore windfarms Dr Jeremy Nedwell, Mr John Langworthy and Mr Daren Howell BWEA Conference 4/3/04 Subacoustech reference: 544R0503, COWRIE Copyright. Aim of study To evaluate the
Search for the missing sea otters
 Search for the missing sea otters Objectives: Based on a real study of sea otter declines Apply your knowledge of basic concepts of population and community ecology Interpret data from graphs and tables
Search for the missing sea otters Objectives: Based on a real study of sea otter declines Apply your knowledge of basic concepts of population and community ecology Interpret data from graphs and tables
Map Showing NAFO Management Units
 Map Showing NAFO Management Units Biology Are 6 species of seals in Atlantic Canadian waters, all of which occur in Newfoundland Two Arctic Species (Ringed, Bearded) Two temperate (Grey, Harbour) Two migratory
Map Showing NAFO Management Units Biology Are 6 species of seals in Atlantic Canadian waters, all of which occur in Newfoundland Two Arctic Species (Ringed, Bearded) Two temperate (Grey, Harbour) Two migratory
What s UP in the. Pacific Ocean? Learning Objectives
 What s UP in the Learning Objectives Pacific Ocean? In this module, you will follow a bluefin tuna on a spectacular migratory journey up and down the West Coast of North America and back and forth across
What s UP in the Learning Objectives Pacific Ocean? In this module, you will follow a bluefin tuna on a spectacular migratory journey up and down the West Coast of North America and back and forth across
Right Whale Diving and Foraging Behavior in the Southwestern Gulf of Maine
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Right Whale Diving and Foraging Behavior in the Southwestern Gulf of Maine Mark Baumgartner Woods Hole Oceanographic Institution
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Right Whale Diving and Foraging Behavior in the Southwestern Gulf of Maine Mark Baumgartner Woods Hole Oceanographic Institution
Blue crab ecology and exploitation in a changing climate.
 STAC Workshop 28 March 2017 Blue crab ecology and exploitation in a changing climate. Thomas Miller Chesapeake Biological Laboratory University of Maryland Center for Environmental Science Solomons, MD
STAC Workshop 28 March 2017 Blue crab ecology and exploitation in a changing climate. Thomas Miller Chesapeake Biological Laboratory University of Maryland Center for Environmental Science Solomons, MD
Acoustic Seaglider for Beaked Whale Detection
 DISTRIBUTION STATEMENT A: Approved for public release; distribution is unlimited. Acoustic Seaglider for Beaked Whale ion Neil M. Bogue and Jim Luby Applied Physics Laboratory University of Washington
DISTRIBUTION STATEMENT A: Approved for public release; distribution is unlimited. Acoustic Seaglider for Beaked Whale ion Neil M. Bogue and Jim Luby Applied Physics Laboratory University of Washington
WHALES. & Whale Sharks
 WHALES & Whale Sharks Introduction Today we are going to learn about the biggest mammals of the ocean and one animal that is commonly mistaken for a whale but is actually a fish. Whale is the common name
WHALES & Whale Sharks Introduction Today we are going to learn about the biggest mammals of the ocean and one animal that is commonly mistaken for a whale but is actually a fish. Whale is the common name
GPS/TDR Satellite Tracking of Sperm Whales with 3-axis Accelerometers- Background. Prepared by Oregon State University.
 GPS/TDR Satellite Tracking of Sperm Whales with 3-axis Accelerometers- Background Prepared by Oregon State University www.soundandmarinelife.org Final Report from Oregon State University's for JIP-sponsored
GPS/TDR Satellite Tracking of Sperm Whales with 3-axis Accelerometers- Background Prepared by Oregon State University www.soundandmarinelife.org Final Report from Oregon State University's for JIP-sponsored
MEASUREMENT OF LONG-TERM AMBIENT NOISE AND TIDAL TURBINE LEVELS IN THE BAY OF FUNDY
 MEASUREMENT OF LONG-TERM AMBIENT NOISE AND TIDAL TURBINE LEVELS IN THE BAY OF FUNDY 11 th European Conference on Underwater Acoustics, 3 July 2012 Bruce Martin 1, Andrew Gerber 2, Christopher Whitt 1,
MEASUREMENT OF LONG-TERM AMBIENT NOISE AND TIDAL TURBINE LEVELS IN THE BAY OF FUNDY 11 th European Conference on Underwater Acoustics, 3 July 2012 Bruce Martin 1, Andrew Gerber 2, Christopher Whitt 1,
FINAL REPORT Tagging and Playback Studies to Toothed Whales
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. FINAL REPORT Tagging and Playback Studies to Toothed Whales Peter Tyack Senior Scientist Emeritus Biology Department Woods
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. FINAL REPORT Tagging and Playback Studies to Toothed Whales Peter Tyack Senior Scientist Emeritus Biology Department Woods
Introduction to Underwater Acoustics Simulator. Poul Kronborg Product Area Owner, Marine MIKE Software Products DHI
 Introduction to Underwater Acoustics Simulator Poul Kronborg Product Area Owner, Marine MIKE Software Products DHI pok@dhigroup.com Module Overview for Marine MIKE Software: Release 2014 Hydrodynamics
Introduction to Underwater Acoustics Simulator Poul Kronborg Product Area Owner, Marine MIKE Software Products DHI pok@dhigroup.com Module Overview for Marine MIKE Software: Release 2014 Hydrodynamics
Blue Whale Behavioral Response Study & Field Testing of the New Bioacoustic Probe
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Blue Whale Behavioral Response Study & Field Testing of the New Bioacoustic Probe Erin M. Oleson NOAA-NMFS-Pacific Islands
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Blue Whale Behavioral Response Study & Field Testing of the New Bioacoustic Probe Erin M. Oleson NOAA-NMFS-Pacific Islands
SEA GRANT PROGRESS REPORT
 SEA GRANT PROGRESS REPORT Project Title: The relationship between seasonal migrations of berried female lobster Homarus americanus, egg development and larval survival. Principal Investigator: Winsor H.
SEA GRANT PROGRESS REPORT Project Title: The relationship between seasonal migrations of berried female lobster Homarus americanus, egg development and larval survival. Principal Investigator: Winsor H.
Alexandre GANNIER. Groupe de Recherche sur les Cétacés, BP 715, Antibes cedex, France. Corresponding author:
 Sci. Rep. Port-Cros natl. Park, 29: 127-134 (2015) Cuvier s beaked whale (Ziphius cavirostris) diving behavior as obtained by visual observation methods and consequences in terms of visual detection during
Sci. Rep. Port-Cros natl. Park, 29: 127-134 (2015) Cuvier s beaked whale (Ziphius cavirostris) diving behavior as obtained by visual observation methods and consequences in terms of visual detection during
Evaluation of fisheries sonar s for whale detection in relation to seismic survey operations
 Evaluation of fisheries sonar s for whale detection in relation to seismic survey operations Simrad AS Norwegian Defense Research Establishment Institute of Marine Research 2 Preface The International
Evaluation of fisheries sonar s for whale detection in relation to seismic survey operations Simrad AS Norwegian Defense Research Establishment Institute of Marine Research 2 Preface The International
Benefit of ESTCP/SERDP Research Program on Underwater Explosive Safety
 Benefit of ESTCP/SERDP Research Program on Underwater Explosive Safety Timothy W. Shelton, PE Supervisory Research Hydraulic Engineer ERDC CHL Oct 2017 DoD has very few options for disposing UXO recovered
Benefit of ESTCP/SERDP Research Program on Underwater Explosive Safety Timothy W. Shelton, PE Supervisory Research Hydraulic Engineer ERDC CHL Oct 2017 DoD has very few options for disposing UXO recovered
Underwater noise The other pollution
 Rebecca Schneider Dept. Natural Resources, Cornell Univ. Underwater noise The other pollution https://i.ytimg.com/vi/ikwqx31mans/maxresdefault.jpg Light decreases with depth in lakes http://academic.keystone.edu/jskinner/limnology/light_in_lakes_lecturenotes.htm
Rebecca Schneider Dept. Natural Resources, Cornell Univ. Underwater noise The other pollution https://i.ytimg.com/vi/ikwqx31mans/maxresdefault.jpg Light decreases with depth in lakes http://academic.keystone.edu/jskinner/limnology/light_in_lakes_lecturenotes.htm
Sound and Light in the Ocean
 Sound and Light in the Ocean When we think of waves in the ocean we often don t think about the sound and light waves that allow for communication and visibility underwater. The underwater environment
Sound and Light in the Ocean When we think of waves in the ocean we often don t think about the sound and light waves that allow for communication and visibility underwater. The underwater environment
Fin Whale. Appendix A: Marine Wildlife. Balaenoptera physalus. New Hampshire Wildlife Action Plan Appendix A Marine-38. Federal Listing.
 Fin Whale Balaenoptera physalus Federal Listing State Listing Global Rank Rank Regional Status E not tracked G3G4TNR State Very High Photo by Christin Khan, NOAA/NEFSC Justification (Reason for Concern
Fin Whale Balaenoptera physalus Federal Listing State Listing Global Rank Rank Regional Status E not tracked G3G4TNR State Very High Photo by Christin Khan, NOAA/NEFSC Justification (Reason for Concern
OVERARCHING PRINCIPLES AND BEST PRACTICE GUIDELINES FOR MARINE MAMMAL WATCHING IN THE WIDER CARIBBEAN REGION (WCR)
") OVERARCHING PRINCIPLES AND BEST PRACTICE GUIDELINES FOR MARINE MAMMAL WATCHING IN THE WIDER CARIBBEAN REGION (WCR) OVERARCHING PRINCIPLES AND BEST PRACTICE GUIDELINES FOR MARINE MAMMAL WATCHING IN THE
OVERARCHING PRINCIPLES AND BEST PRACTICE GUIDELINES FOR MARINE MAMMAL WATCHING IN THE WIDER CARIBBEAN REGION (WCR) OVERARCHING PRINCIPLES AND BEST PRACTICE GUIDELINES FOR MARINE MAMMAL WATCHING IN THE
Issues Affecting the Southern Resident Orcas
 Reading Practice A Issues Affecting the Southern Resident Orcas Orcas, also known as killer whales, are opportunistic feeders, which means they will take a variety of different prey species. J, K, and
Reading Practice A Issues Affecting the Southern Resident Orcas Orcas, also known as killer whales, are opportunistic feeders, which means they will take a variety of different prey species. J, K, and
Characterizing The Surf Zone With Ambient Noise Measurements
 Characterizing The Surf Zone With Ambient Noise Measurements LONG-TERM GOAL Grant Deane Marine Physical Laboratory Scripps Institution of Oceanography La Jolla, CA 93093-0213 phone: (619) 534-0536 fax:
Characterizing The Surf Zone With Ambient Noise Measurements LONG-TERM GOAL Grant Deane Marine Physical Laboratory Scripps Institution of Oceanography La Jolla, CA 93093-0213 phone: (619) 534-0536 fax:
Daniel E. Crocker Department of Biology, Sonoma State University, Rohnert Park, California
 The effect of a low-frequency sound source (acoustic thermometry of the ocean climate) on the diving behavior of juvenile northern elephant seals, Mirounga angustirostris Daniel P. Costa a) Department
The effect of a low-frequency sound source (acoustic thermometry of the ocean climate) on the diving behavior of juvenile northern elephant seals, Mirounga angustirostris Daniel P. Costa a) Department
Marine Mammals in Faroese Waters
 Marine Mammals in Faroese Waters With Special Attention to the South-south-eastern Sector of the Region by Dorete Bloch 1, Bjarni Mikkelsen 2 and Lise H. Ofstad 2 1 Museum of Natural History 2 Faroese
Marine Mammals in Faroese Waters With Special Attention to the South-south-eastern Sector of the Region by Dorete Bloch 1, Bjarni Mikkelsen 2 and Lise H. Ofstad 2 1 Museum of Natural History 2 Faroese
Wave Prediction in the Santa Barbara Channel
 Wave Prediction in the Santa Barbara Channel OCS Study MMS 2001-055 Final Technical Summary Final Study Report U.S. Department of the Interior Minerals Management Service Pacific OCS Region Wave Prediction
Wave Prediction in the Santa Barbara Channel OCS Study MMS 2001-055 Final Technical Summary Final Study Report U.S. Department of the Interior Minerals Management Service Pacific OCS Region Wave Prediction
Nancy E. Kohler, Danielle Bailey, Patricia A. Turner, and Camilla McCandless SEDAR34-WP-25. Submitted: 10 June 2013
 Mark/Recapture Data for the Atlantic Sharpnose Shark (Rhizoprionodon terranovae), in the Western North Atlantic from the NEFSC Cooperative Shark Tagging Program Nancy E. Kohler, Danielle Bailey, Patricia
Mark/Recapture Data for the Atlantic Sharpnose Shark (Rhizoprionodon terranovae), in the Western North Atlantic from the NEFSC Cooperative Shark Tagging Program Nancy E. Kohler, Danielle Bailey, Patricia
Understanding the Foraging Ecology of Beaked and Short-Finned Pilot Whales in Hawaiian Waters
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Understanding the Foraging Ecology of Beaked and Short-Finned Pilot Whales in Hawaiian Waters PI: Whitlow W. L. Au 1 CoPI:
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Understanding the Foraging Ecology of Beaked and Short-Finned Pilot Whales in Hawaiian Waters PI: Whitlow W. L. Au 1 CoPI:
Modeling Gas Dynamics in California Sea Lions
 DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Modeling Gas Dynamics in California Sea Lions Andreas Fahlman Department of Life Sciences Texas A&M University-Corpus Christi
DISTRIBUTION STATEMENT A. Approved for public release; distribution is unlimited. Modeling Gas Dynamics in California Sea Lions Andreas Fahlman Department of Life Sciences Texas A&M University-Corpus Christi
Modern status of Sea otter population on the Commander Islands
 Modern status of Sea otter population on the Commander Islands Alexander Burdin, Kamchatka branch of Pacific Institute of Geography, RAS, University of Alaska, Fairbanks, ASLC, Sergey Zagrebelny, Commander
Modern status of Sea otter population on the Commander Islands Alexander Burdin, Kamchatka branch of Pacific Institute of Geography, RAS, University of Alaska, Fairbanks, ASLC, Sergey Zagrebelny, Commander
RESOLUTION 4.17 GUIDELINES TO ADDRESS THE IMPACT OF ANTHROPOGENIC NOISE ON CETACEANS IN THE ACCOBAMS AREA
 RESOLUTION 4.17 GUIDELINES TO ADDRESS THE IMPACT OF ANTHROPOGENIC NOISE ON CETACEANS IN THE ACCOBAMS AREA The Meeting of the Parties to the Agreement on the Conservation of Cetaceans of the Black Sea,
RESOLUTION 4.17 GUIDELINES TO ADDRESS THE IMPACT OF ANTHROPOGENIC NOISE ON CETACEANS IN THE ACCOBAMS AREA The Meeting of the Parties to the Agreement on the Conservation of Cetaceans of the Black Sea,
Introduction. o 2. ! "#$ % & ' (" 4 Watt/m 2. Major
 07, 08 9 07%, 8 Abstract Killer whale pods sometimes hunt herring by corralling the fish into a tight ball near the ocean surface and stunning them with underwater tail slaps before eating them. I asked
07, 08 9 07%, 8 Abstract Killer whale pods sometimes hunt herring by corralling the fish into a tight ball near the ocean surface and stunning them with underwater tail slaps before eating them. I asked
MANAGEMENT OF KRILL AS AN ESSENTIAL COMPONENT OF THE CALIFORNIA CURRENT ECOSYSTEM
 MANAGEMENT OF KRILL AS AN ESSENTIAL COMPONENT OF THE CALIFORNIA CURRENT ECOSYSTEM AMENDMENT 12 TO THE COASTAL PELAGIC SPECIES FISHERY MANAGEMENT PLAN ENVIRONMENTAL ASSESSMENT, REGULATORY IMPACT REVIEW
MANAGEMENT OF KRILL AS AN ESSENTIAL COMPONENT OF THE CALIFORNIA CURRENT ECOSYSTEM AMENDMENT 12 TO THE COASTAL PELAGIC SPECIES FISHERY MANAGEMENT PLAN ENVIRONMENTAL ASSESSMENT, REGULATORY IMPACT REVIEW
Behavioral Response of Dolphins to Signals Simulating Mid-Frequency Sonar
 DISTRIBUTION STATEMENT A: Approved for public release; distribution is unlimited. Behavioral Response of Dolphins to Signals Simulating Mid-Frequency Sonar Dorian S. Houser Biomimetica 7951 Shantung Drive
DISTRIBUTION STATEMENT A: Approved for public release; distribution is unlimited. Behavioral Response of Dolphins to Signals Simulating Mid-Frequency Sonar Dorian S. Houser Biomimetica 7951 Shantung Drive
Can we be as clever as a sperm whale?
 Can we be as clever as a sperm whale? { Efficacy of Counter-Measures to Reduce Sperm Whale Depredation on Demersal Longlines in the Gulf of Alaska Wild, Straley, Thode, Liddle, Mathias, O Connell, Behnken,
Can we be as clever as a sperm whale? { Efficacy of Counter-Measures to Reduce Sperm Whale Depredation on Demersal Longlines in the Gulf of Alaska Wild, Straley, Thode, Liddle, Mathias, O Connell, Behnken,
Killer whales and their prey in Iceland
 Killer whales and their prey in Iceland Filipa I. P. Samarra Marine and Freshwater Research Institute, Skúlagata 4, 101 Reykjavík, Iceland June-August 2018 PAGE 1 Letter to Volunteers Dear Earthwatch volunteers,
Killer whales and their prey in Iceland Filipa I. P. Samarra Marine and Freshwater Research Institute, Skúlagata 4, 101 Reykjavík, Iceland June-August 2018 PAGE 1 Letter to Volunteers Dear Earthwatch volunteers,
Tracking Juvenile Summer Flounder
 Tracking Juvenile Summer Flounder East Coast MARE Materials For the leader: Whiteboard Markers (different colors) For each group: Copies of student group packets Copies of student worksheet Overview Scientists
Tracking Juvenile Summer Flounder East Coast MARE Materials For the leader: Whiteboard Markers (different colors) For each group: Copies of student group packets Copies of student worksheet Overview Scientists
Unalaska Navigation Channel Improvements
 1 237 237 237 255 255 255 0 0 0 Unalaska Navigation Channel Improvements 217 217 217 200 200 200 163 163 163 131 132 122 239 65 53 80 119 27 252 174.59 110 135 120 112 92 56 62 102 130 102 56 48 130 120
1 237 237 237 255 255 255 0 0 0 Unalaska Navigation Channel Improvements 217 217 217 200 200 200 163 163 163 131 132 122 239 65 53 80 119 27 252 174.59 110 135 120 112 92 56 62 102 130 102 56 48 130 120
ATLANTIC STURGEON. Consultations on listing under the Species at Risk Act
 ATLANTIC STURGEON Consultations on listing under the Species at Risk Act Information summary and questionnaire for consultations on adding two Atlantic Sturgeon populations to the List of Wildlife Species
ATLANTIC STURGEON Consultations on listing under the Species at Risk Act Information summary and questionnaire for consultations on adding two Atlantic Sturgeon populations to the List of Wildlife Species
BEFORE YOU OPEN ANY FILES:
 Dive Analysis Lab * Make sure to download all the data files for the lab onto your computer. * Bring your computer to lab. * Bring a blank disk or memory stick to class to save your work and files. The
Dive Analysis Lab * Make sure to download all the data files for the lab onto your computer. * Bring your computer to lab. * Bring a blank disk or memory stick to class to save your work and files. The
Observing the behavioral response of herring exposed to mid-frequency sonar signals
 Observing the behavioral response of herring exposed to mid-frequency sonar signals Handegard 1, Nils Olav and Doksaeter 1, Lise and Godoe 1, Olav Rune and Kvadsheim 2, Petter H. 1 Institute of Marine
Observing the behavioral response of herring exposed to mid-frequency sonar signals Handegard 1, Nils Olav and Doksaeter 1, Lise and Godoe 1, Olav Rune and Kvadsheim 2, Petter H. 1 Institute of Marine
Caltrans compendium of underwater sound data from pile driving 2014 update
 Caltrans compendium of underwater sound data from pile driving 2014 update Richard RODKIN 1 ; Keith POMMERENCK 2 1 Illingworth & Rodkin, Inc., United States 2 Illingworth & Rodkin, Inc., United States
Caltrans compendium of underwater sound data from pile driving 2014 update Richard RODKIN 1 ; Keith POMMERENCK 2 1 Illingworth & Rodkin, Inc., United States 2 Illingworth & Rodkin, Inc., United States
Climate Change and Arctic Marine Mammals: Living on the Edge
 Climate Change and Arctic Marine Mammals: Living on the Edge M.O. Hammill Maurice Lamontagne Institute Dept. of Fisheries & Oceans Canada Mont-Joli, QC, Canada Outline: Hudson Bay- Bears (losers?) Killer
Climate Change and Arctic Marine Mammals: Living on the Edge M.O. Hammill Maurice Lamontagne Institute Dept. of Fisheries & Oceans Canada Mont-Joli, QC, Canada Outline: Hudson Bay- Bears (losers?) Killer
no-take zone 1 of 5 Channel Islands National Marine Sanctuary, California
 This website would like to remind you: Your browser (Apple Safari 4) is out of date. Update your browser for more security, comfort and the best experience on this site. Encyclopedic Entry no-take zone
This website would like to remind you: Your browser (Apple Safari 4) is out of date. Update your browser for more security, comfort and the best experience on this site. Encyclopedic Entry no-take zone
Underwater Behavior of Blue Whales Examined with Suction-cup Attached Tags
 Underwater Behavior of Blue Whales Examined with Suction-cup Attached Tags John Calambokidis Cascadia Research 218½ 4 th Ave. Olympia, WA 98501 Phone: (360) 943-7325 fax: (360)943-7026 email: Calambokidis@cascadiaresearch.org
Underwater Behavior of Blue Whales Examined with Suction-cup Attached Tags John Calambokidis Cascadia Research 218½ 4 th Ave. Olympia, WA 98501 Phone: (360) 943-7325 fax: (360)943-7026 email: Calambokidis@cascadiaresearch.org