THE INFLUENCE OF TEMPERATURE ON MUSCLE VELOCITY AND SUSTAINED PERFORMANCE IN SWIMMING CARP
|
|
|
- Adela Morrison
- 5 years ago
- Views:
Transcription
1 J. exp. Biol. 154, (1990) 163 Printed in Great Britain The Company of Biologists Limited 1990 THE INFLUENCE OF TEMPERATURE ON MUSCLE VELOCITY AND SUSTAINED PERFORMANCE IN SWIMMING CARP BY LAWRENCE C. ROME, ROEL P. FUNKE* AND R. McNEILL ALEXANDERt Department of Biology, Leidy Laboratories, University of Pennsylvania, Philadelphia, PA 19104, USA Accepted 14 June 1990 Summary The aim of this study was to evaluate how fish locomote at different muscle temperatures. Sarcomere length excursion and muscle shortening velocity, V, were determined from high-speed motion pictures of carp, Cyprinus carpio (11-14cm), swimming steadily at various sustained speeds at 10, 15 and 20 C. In the middle and posterior regions of the carp, sarcomeres of the lateral red muscle underwent cyclical excursions of 0.31 fim, centred around the resting length of 2.06/mi (i.e. from 1.91 to 2.22^m). The amplitudes of the sarcomere length excursions were essentially independent of swimming speed and temperature. As tail-beat frequency increased linearly with swimming speed regardless of temperature, the sarcomeres underwent the same length changes in a shorter time. Thus, V increased in a linear and temperature-independent manner with swimming speed. Neither temperature nor swimming speed had an influence on tail-beat amplitude or tail height. Our findings indicate that muscle fibres are used only over a narrow, temperature-independent range of V/V max ( ) where power and efficiency are maximal. Carp start to recruit their white muscles at swimming speeds where the red muscle V/V max becomes too high (and thus power output declines). When the V/K max of the active muscle falls too low during steady swimming, carp switch to 'burst-and-coast' swimming, apparently to keep V/V mak high. Because K max (maximum velocity of shortening) of carp red muscle has a Qio of 1.63, the transition speeds between swimming styles are lower at lower temperatures. Thus, carp recruit their white anaerobic muscle at a lower swimming speed at lower temperatures (verified by electromyography), resulting in a lower maximum sustainable swimming speed. The present findings also indicate that, to generate the same total force and power to swim at a given speed, * Present address: Department of Plant Molecular Biology, University of Tennessee, Knoxville, TN 37996, USA. t Present address: Department of Pure and Applied Biology, University of Leeds, Leeds LS2 9JT, England. Key words: muscle, sarcomere length, swimming, Cyprinits carpio.
 of vertebrate skeletal muscle has a Q 10 value of")
2 164 L. C. ROME, R. P. FUNKE AND R. MCN. ALEXANDER carp at 10 C must recruit about 50% greater fibre cross-sectional area than they do at 20 C. Introduction Temperature has a strong influence on the contractile properties of muscles: for instance, the maximum velocity of shortening (V max ) of vertebrate skeletal muscle has a Q 10 value of approximately (Bennett, 1984). Most ectothermal animals are subjected to considerable acute, diurnal and seasonal changes in body temperature, and it would appear from the thermal sensitivity of their muscle V max that they would be faced with significant problems when trying to locomote at different temperatures. Despite changes in the contractile properties of their muscles, ectotherms are able to locomote over a wide range of temperatures. Further, the thermal dependence of the contractile properties seems to have no apparent effect on the kinematics of animals locomoting over a moderate range of sustainable speeds (Rome, 1982; Rome etal. 1984). To locomote at a given speed the muscle must generate the same force and mechanical power irrespective of temperature. Owing to the change in V max, cold muscle is unable to generate as much force and power per active cross-section as warm muscle. Based on electromyographic experiments on carp, Rome and colleagues (Rome etal. 1984, 1985; Rome, 1986, 1990) hypothesized that to generate the same force and power the animal must recruit more aerobic red fibres at lower temperatures. According to this 'compression of the recruitment order' theory, motor units are recruited in the same order at all temperatures but, to compensate for their reduced power output, the motor units are recruited over a lower range of speeds at lower temperatures. Recruiting all of their aerobic fibres (and hence having to recruit their anaerobic fibres) at a lower speed, however, results in reduced sustainable performance. In a previous study, Rome et al. (1988) observed that important functional parameters of muscle (force production, power production and efficiency) at a given temperature depend on V/V mak, where Fis the velocity at which the fibres shorten. In this paper, we measure the influence of temperature on V of the lateral red muscle at various swimming speeds in carp. Rome and Sosnicki (1990) previously determined the influence of temperature on V max of carp red muscle. Thus, with values for V/V max during swimming at different temperatures we can gain further insight into the problems faced and the solutions taken by ectotherms locomoting with different muscle temperatures. Materials and methods Animals Carp (Cyprinus carpio L.) were purchased from a commercial fishery and kept in a tank at a water temperature of 15 C for 6 weeks prior to these experiments.

3 Muscle velocity in swimming carp 165 The fish (ll-14cm in length) were fed daily ad libitum and kept on a 14h:10h light:dark cycle. Measurement of sarcomere length during swimming Our goal was to measure the sarcomere length changes with time that red muscle undergoes during swimming. The methods used for these procedures are carefully described elsewhere (L. C. Rome and A. A. Sosnicki, in preparation). Briefly, sarcomere length (SL) of the red muscle can be described by the following equations: SL concave = A/R (r 2 = 0.98), where R is the radius of curvature at the backbone, and A is the half-thickness of the fish. This relationship was derived empirically by killing fish, bending them into shapes obtained during swimming, allowing them to go into rigor, and then measuring sarcomere length as a function oia/r in frozen sections. Note that in a straight fish, R is infinite, and thus sarcomere length is 2.06(xm. Having established this relationship, sarcomere length of the lateral red muscle was calculated from the changes of curvature of fishes' bodies during the tail-beat cycle. This involved digitizing the outline of the fish and determining A/R, as in Rome etal. (1988). Carp were filmed from above in a recirculating water treadmill. The water treadmill consisted of a variable-drive motor, centrifugal pump, plastic pipe sections and an acrylic swim tube (diameter 16 cm). The temperature of the unit was controlled to within 1 C, and the fish were confined to the swim tube with upstream and downstream honeycomb grids. The flow velocity and velocity profile of the water treadmill were measured with a Swoffer water velocity meter. Although velocity dropped by about 5 cm s" 1 within 3 cm of the walls of the swim tube, velocity in the central 10 cm of the section was quite uniform and steady with time. Four carp were filmed (200 frames s" 1 ) from above swimming at speeds ranging from 15 to 45cms" 1 with a high-speed motion picture camera (LOCAM, Redlake Corp.). To obtain a sharp silhouette of the fish in very low light levels, the bottom of the swim tube was covered with reflector material ( Scotchlite, as in Wardle, 1975) and illuminated with a ring light mounted on the camera lens. Rome et al. (1984,1985) found that carp avoid bright lights and prefer to swim in areas of darkness. Thus, low light levels were used and the sides and back of the swim tube were illuminated with narrow-beam high-intensity lights to prevent the fish from drifting back or taking advantage of decreased flow rates near the walls. These precautions, and the fact that the fish were too large to benefit much from the Ireduced flow rate near the walls of the flume, ensured that they were swimming against the full force of the water current. Following the first series of swims at 15 C (see Rome et al. 1988), fish swam at 10
 prior to experimentation.")
4 166 L. C. ROME, R. P. FUNKE AND R. MCN. ALEXANDER and 20 C, with at least 48 h between bouts of exercise. Animals were held at the new swimming temperature for 24 h before filming. Fish were placed in the swim tube and allowed to adjust for at least 15min with the water circulating at a moderate speed (about 15cms~ ] ) prior to experimentation. The criteria for steady swimming at a certain speed were that the fish should be swimming steadily in the middle of the flume with minimal acceleration or deceleration for at least five complete tail-beat cycles. Usually three tail-beat cycles were analyzed (see Fig. 2). At slow speeds (15-25cms" 1 ) sufficient resolution was achieved by analyzing every fourth or fifth frame of a sequence, whereas at higher speeds it was necessary to analyze every third or second frame. Thus, depending on swimming speed, each sequence contained frames. Sarcomere lengths on the concave and convex sides were calculated for three positions along the length of the fish: 38 %, 52 % and 68 % of the overall length, and labelled anterior, middle and posterior, respectively. The plots of sarcomere length against time appeared to be best described as triangular waves. The velocity of shortening of the muscle was estimated for the three positions on the fish at each swimming speed using a five-point smoothing and differentiation scheme (Lanczos, 1956). The highest velocities were obtained in the rising and falling parts of the waves, and the lowest velocities at the crests and troughs, where the slope was changing sign. Sufficiently small frame intervals at each swimming speed were chosen so that the five-point smoothing and differentiation scheme gave accurate values for the maximal velocity (i.e. the endpoints did not lie in the crests and troughs of the graph). Because the rising part of the curve represented shortening of sarcomeres on the right side of the fish and the falling part of the curve represented shortening of the sarcomeres on the left side, the maximum slopes on the rising and falling portions of the curve were taken as shortening velocities. V was thus calculated as the average of the five or six maximal velocities obtained from three tail beats in each sequence. The contribution of the tendons in the red muscle to the overall length change was considered to be negligible. In fish, the red muscle fibres are attached end to end through short myosepta. These myosepta are thin and their length change would not represent an appreciable percentage of the total change in length of the muscle-tendon complex. Measurement of tail-beat amplitude and tail height For each of the sequences used to determine V, the amplitude of the tail beat was also measured. Generally, at least 10 tail-beat cycles were examined and excursions averaged. In addition, the fish were filmed from the side at each speed to look at fluctuations in tail height during swimming. A Bolex motion picture camera (framing rate 64 s" 1 ) was placed lm from the swim tube using the reflector material ( Scotchlite) as a backdrop. With the fish swimming steadily in middle of the tube, the error due to parallax was about 5 %, and the measurement were adjusted accordingly. We found the same rhythmical fluctuations in height as
5 Muscle velocity in swimming carp 167 o.u 5.5 I 50 J2 4.0 ( #< / >m ' *\ a«mean= #/ ' ' cm 0. oo Time (s) 0.75 l.( Fig. 1. Representative record of fluctuating tail height during two tail-beat cycles in a carp swimming at 20 cms" 1. Measurements were taken at the trailing edge of the tail, and mean values were used to test the influence of swimming speed and temperature on tail height. Bainbridge (1963), and used an average value calculated over about 8-10 tail beats (Fig. 1). Electromyography The swimming speed at which white muscle recruitment occurred was determined at each temperature by monitoring the activity of the white muscle by electromyography (see Rome et al. 1984, 1985). Electrodes were implanted in the white muscle on both sides of the fish with a hypodermic needle (the activity of red muscle has been previously studied in considerable detail; Rome et al. 1984,1985). During this procedure fish were held in a surgical rack and anaesthetized by perfusing their gills with SOmgP 1 tricaine (MS222). The electrodes were attached to high-impedance probes. Signals from the muscle were passed through a.c. preamplifiers (Grass no. P511), and recorded on two audio tracks (Cannon) of simultaneous video tape of the fish swimming during the electromyography experiments. The time and swimming speed were placed on the tape with a time-date generator (Panasonic) and video typewriter (Fora VT-100). We found that the electrodes picked up electrical spikes from the motor of the swimming machine. Electrical interference was eliminated by building a Faraday cage around the swimming tube and enclosing the high-impedance probes within it. Typically, there was only 10-20;uV of noise. The flow velocity was increased in steps of 5cms" 1 (smaller increments of 2.5cms"" 1 were used near the recruitment speed), and the recruitment speed was taken to be halfway between the lowest speed at which the fish was unable to fcnaintain position without regularly using its white muscle and the speed immediately below it. Recruitment speeds at all three temperatures were determined in the same

6 168 L. C. ROME, R. P. FUNKE AND R. MCN. ALEXANDER animals subsequent to the filming. Recruitment speeds at 10 and 20 C were additionally determined on carp that were not filmed. Results Influence of temperature on sarcomere length excursion and velocity of shortening Sarcomere length during swimming was determined from the curvature of the fishes' backbone in the films. Fig. 2 shows a typical record of the changes in sarcomere length at three places along the body of a fish swimming at 25 and at 40 cms" 1. In general, the anterior position was noisier than the middle and posterior positions. This is because curvatures were smaller in that position, and random errors (e.g. due to tracing the side of the fish with a cursor) are more apparent. In addition, the pelvic fins sometimes interrupted the profile of the side of the fish and we had to make an approximation by drawing through them. This noise made it difficult to pick out the waveform of sarcomere length with time (see especially the first half of the trace in Fig. 2B). It was thus not possible to measure accurately the amplitude (sarcomere length excursion) of the anterior waveform or its slope (muscle shortening velocity). As swimming speed and sarcomere length excursion increased, the signal-to-noise ratio in the anterior position improved, as can be seen at 40cms" 1 (Fig. 2C). We were able to film some of the fish swimming steadily at speeds ranging from 15-30, and 20-45cms"" 1 at 10, 15 and 20 C, respectively (some carp could swim to only 5cms" 1 below the maximal speeds quoted above; also 15 C values are recalculated from Rome etal. 1988). The maximum speeds at which the carp were able to swim steadily at each temperature correlated well with the initial speeds of white muscle recruitment determined by electromyography (27.1 cms" 1 ±S.E. = 1.8, N=8 at 10 C, 36.30ns- 1 ±S.E. = 1.4, N=8 at 15 C and 42.9cms-' ±S.E.=3.5, N=5 at 20 C). Thus, at the swimming speeds we filmed, the carp were using their red muscle fibres exclusively. At higher speeds at each temperature the white muscle was recruited and the fish switched to a burst-and-coast mode of swimming. At swimming speeds below the minimum for steady swimming, the fish also used the burst-and-coast mode of swimming, but this involved recruitment of the red muscle exclusively. For instance, the fish would not swim steadily at 15cms" 1 at 15 or 20 C, whereas they did so at 10 C. Fig. 3 shows sarcomere length excursion in the middle and posterior positions at all three experimental temperatures over the range of speeds at which the fish swam steadily. Excursions ranged from 0.21 to 0.31 ^m, centred around the sarcomere length in a straight fish of 2.06^m. In these positions the excursions were essentially independent of swimming speed and temperature. The excursion^ in the anterior position were undoubtedly smaller (i.e. making them unanalyzable) than those in the middle and posterior positions at low swimming speeds, but they
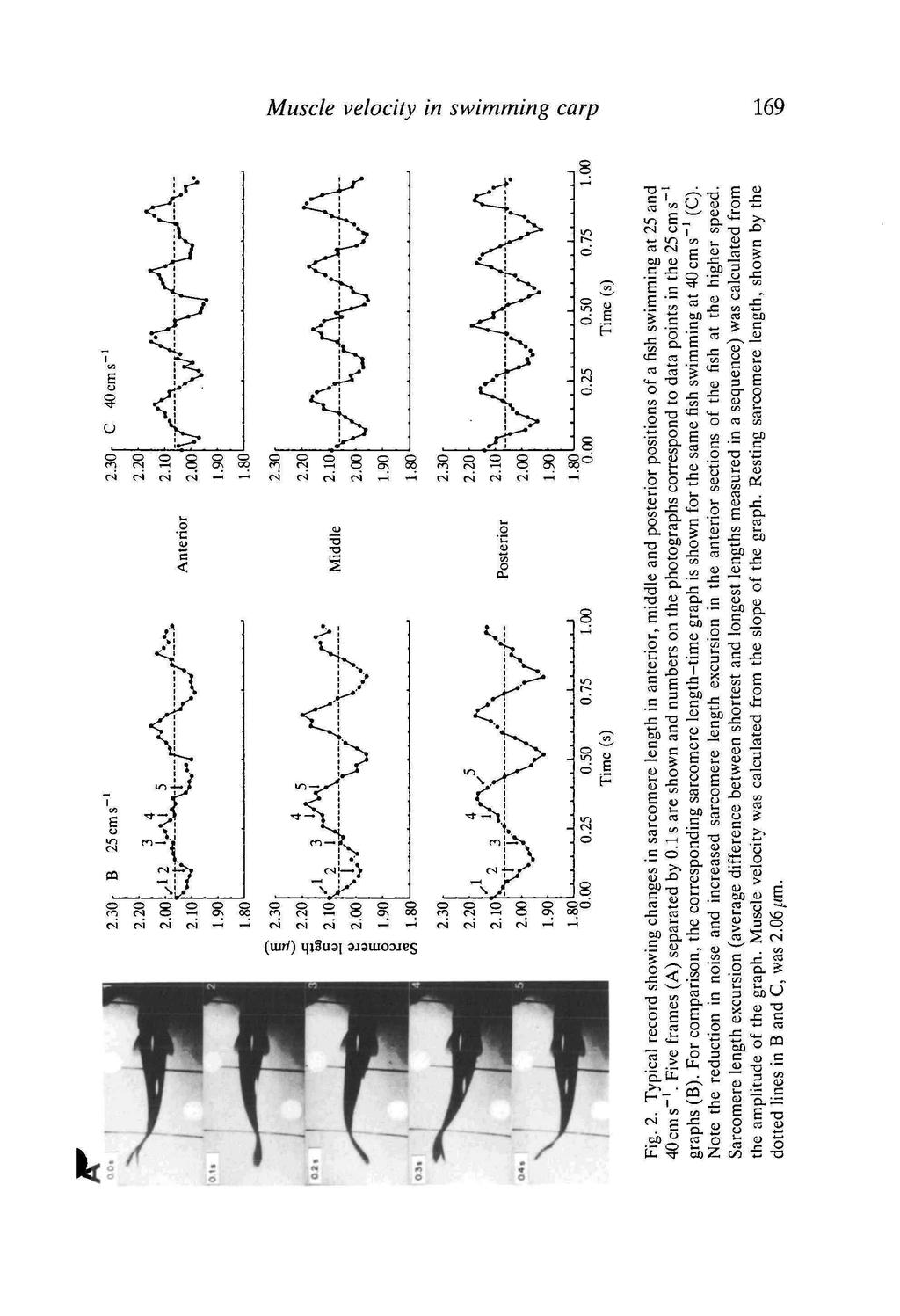
7
8 170 L. C. ROME, R. P. FUNKE AND R. MCN. ALEXANDER 0.4 Middle ' 0.1 n n i j., 1 a ft n A r ii, i 10 C D 15 C A 20 C Swimming speed (cms" 1 ) Fig. 3. Relationship between swimming speed and sarcomere length excursion at the three experimental temperatures in the middle and posterior positions on the fish. Points are means±s.e. (N=2-5). The influence of swimming speed in the middle and posterior positions was small (only at 15 C in the middle is the linear regression significant at the 0.05 confidence level). Further, temperature had no effect on sarcomere length excursion in the middle and posterior positions. increased more significantly as the fish swam faster (i.e. the waveform became more distinct). Tail-beat frequency was found to increase linearly with swimming speed and was independent of temperature (Fig. 4). Thus, the sarcomeres underwent the same length changes in a shorter period, resulting in an increase in muscle shortening velocity, as is shown in Fig. 5. Fig. 5 also shows that temperature had no effect on the velocity of shortening of the red muscle in the middle and posterior positions. Thus, the movements of the muscle fibres at a given swimming speed are independent of temperature. Two parameters that influence the amount of thrust transmitted to the water are tail-beat amplitude and tail height. Fig. 6 shows that neither temperature nor swimming speed had an effect on tail-beat amplitude or tail height. Discussion Influence of temperature on kinematics of steady swimming and sustained performance Over the range of steady swimming speeds, there were no changes in tail-beat amplitude or tail height with swimming speed. These results suggest that the
9 Muscle velocity in swimming carp Swimming speed (cms" 1 ) 40 Fig. 4. Temperature had no effect on tail-beat frequency (ANCOVA, P>0.1). Linear increase of tail-beat frequency with swimming speed at all three temperatures is shown by the solid line (frequency=0.096xvelocity+1.17, ^=0.98, N=52). Points are means+s.e Middle C D 15 C A 20 C Posterior Swimming speed (cm s~') Fig. 5. Relationship between swimming speed and muscle velocity at 10, 15 and 20 C in the middle and posterior positions. Muscle velocities increase linearly with swimming speed at both positions (r 2 =0.98 and 0.95 in the middle and posterior positions, respectively, N=53), due to the increase in tail-beat frequency. Temperature had no effect on velocities in the middle and posterior positions (ANCOVA, P>0.25 in both positions). Points are means±s.e.
10 172 L. C. ROME, R. P. FUNKE AND R. MCN. ALEXANDER f » > < I A Q C D 15 C A 20 C o.u (cm) < x: 3.0 op , i B i T V 0 D A T A D 10 C 15 C 20 C I n n 1 i Swimming speed (cm s~ ) i Fig. 6. Neither swimming speed nor temperature had an influence on tail-beat amplitude or tail height (ANOVA, P>0.25). Points are means±s.e. (N=2-5). increased thrust at increasing speeds was provided solely by the linear increase in tail-beat frequency with swimming speed (in agreement with Hunter and Zweiffel, 1971). As sarcomere length excursion was independent of swimming speed, the increase in tail-beat frequency resulted in a linear increase in muscle shortening velocity with increasing swimming speed. None of the above kinematic parameters at a given swimming speed were influenced by temperature. Previous studies of temperature effects on tail-beat frequency have shown similar temperature independence in cyprinids (Smit et al. 1971; Rome etal. 1984) and small temperature dependences in trout Salmo gairdneri, bass Micropterus salmoides (Stevens, 1979) and striped bass Morone saxatilis (Sisson and Sidell, 1987). The temperature independence of kinematic parameters in this study shows that, despite large changes in contractile properties, the nervous system of carp is capable of recruiting muscle so that movement at a given speed is the same at all temperatures. Similar results have been found in a very different locomotory system: lizards Varanus exanthematicus running at different muscle temperatures (Rome, 1982). These results suggest that there is ani optimal way for an animal to locomote in a particular medium, and that this depends on anatomical parameters (e.g. origin and insertion of fibres) and the
![Muscle velocity in swimming carp 173 physical properties of the environment, which are nearly independent of temperature [except for the small increase (10%) in drag of water at low temperatures].](/docs-images/91/106560548/images/11-0.jpg "The mechanism by which animals produce the same movements with the. reduced power output of cold muscle has been termed 'compression of the recruitment order' (Rome etal.")
11 Muscle velocity in swimming carp 173 physical properties of the environment, which are nearly independent of temperature [except for the small increase (10%) in drag of water at low temperatures]. The mechanism by which animals produce the same movements with the. reduced power output of cold muscle has been termed 'compression of the recruitment order' (Rome etal. 1984, 1985; Rome, 1986, 1990). Motor units are recruited in the same order at low and high temperatures (red muscle motor units at low speeds followed by white ones at high speeds), but at low temperatures the order is compressed into a reduced speed range. Thus, at a given speed, more muscle fibres are recruited at the lower temperatures, compensating for their lower relative force and power outputs. This mechanism seems far more likely than the alternative mechanism of pulse coding, whereby the cold muscle compensates by having a higher level of activation (Rome, 1990). Because fish must recruit more muscle fibres as they swim faster at any given temperature, the carp reaches a speed (maximum sustainable speed) at which it has recruited all its aerobic fibres. To generate the additional power needed to swim faster, the white anaerobic fibres must be recruited. Recruitment of the white anaerobic muscle at lower speeds at low temperatures (as observed in this study) provides the strongest evidence for the 'compression of the recruitment order' theory. Early recruitment of white muscle at low temperatures has also been reported in other fish (striped bass; Sisson and Sidell, 1987) and in lizards Varanus exanthematicus (Jayne et al. 1988). The findings in the present study agree almost perfectly with those found in previous studies of white muscle recruitment speeds in carp (26cms" 1 at 10 C and 46cms~ 1 at 20 C; Rome etal. 1984). We observed in this study, as well as in previous studies, that recruitment of white muscle resulted in rapid fatigue. Thus, maximum sustainable swimming speed is reduced by about one-third at 10 C compared to 20 C. V/V maj: : a determinant of locomotory behaviour of carp at different temperatures Like cod Gadus morhua (Videler, 1981) and saithe Pollachius virens (Videler and Weihs, 1982), carp used three distinct phases of swimming: burst-and-coast swimming powered by red muscle at relatively low swimming speeds, steady swimming powered by red muscle at moderate speeds (discussed above) and burstand-coast swimming powered by red and white muscle at high swimming speeds. All three phases were present at 10 and 20 C; however, the swimming speeds at which the transitions occurred between them were different at these two temperatures. Comparison of the value of V at which fibres are shortening during locomotion with the force-velocity characteristics of the fibres provides considerable insight into why these three phases occur and how the transition speeds vary with temperature. Fig. 7 shows the average force-velocity and power-velocity characteristics of carp red muscle at 10 and 20 C based on measurements by Rome and Sosnicki 1(1990). The shaded region on each curve (outlined by vertical dotted lines) represents the range of shortening velocities, V, that were observed in this study during steady swimming. As the relationship between muscle shortening velocity
12 174 ROME, R. P. FUNKE AND R. MCN. ALEXANDER Muscle velocity (MLs ') Swimming speed (cms" 1 ) Fig. 7 and swimming speed was independent of temperature, the swimming speed axis is also shown and power, force and V of the red muscle can be related. In terms of swimming speed, the dotted vertical lines represent the transition points between steady swimming and the two phases of burst-and-coast swimming. At swimming speeds below that of the left-hand dotted line, fish burst-and-coast with red muscle. At swimming speeds above that of the right dotted line, the fish burst-andcoast with their white muscle. Below we focus on steady swimming, followed by a brief discussion of burstand-coast swimming at high and at low speeds. Steady swimming,-i The carp swam steadily between 15 and 30 cms" 1 at 1O C and between 20 and 45cms" 1 at 20 C. If cold fish compensate solely by recruiting more fibres, then
 force-velocity and power-velocity curves of carp red muscle at 10 and 20 C based on the results of Rome and Sosnicki (1990; the dotted portions of the")
13 Muscle velocity in swimming carp 175 Fig. 7. Influence of temperature on mechanical properties of carp red muscle and on their use during swimming. This figure shows the average (see note below) force-velocity and power-velocity curves of carp red muscle at 10 and 20 C based on the results of Rome and Sosnicki (1990; the dotted portions of the curves were not covered by their experiments and are fitted by eye based on previous results). Rome and Sosnicki (1990) found that V max, maximum power and maximum force generation were 3.55MLs~\ eowkg" 1 and 109kNm" 2, respectively, at 10 C and 5.71MLs~\ 94Wkg~' ' and 123kNm" 2, respectively, at 20 C, where ML is muscle length. As muscle shortening velocity during steady swimming was independent of temperature, we have placed the swimming speed axis on the graph as well (note that at 15 and 45cms" 1, the scale is shifted slightly to coincide with the actual values of Vobserved). Thus, during steady swimming, the curves provide the power, force and muscle shortening velocity as a function of swimming speed. The shaded regions represent the muscle shortening velocity during steady swimming with red muscle. The dotted vertical lines at each temperature represent transition swimming speeds. At slower swimming speeds than that of the left-hand line, the carp used burst-and-coast swimming with red muscle. At higher swimming speeds than that of the right-hand line, the white muscle is recruited and the carp used burst-and-coast swimming. For each temperature, the V/ V max at the transition points is given. In addition to the above V max values, the curves were defined by the following coefficients: a/p o *=0.455 and P o * = 1.19 at 10 C and a/p o *=0.30 and P O * = 1.29 at 20 C, where a is the Hill constant and P o * is the extrapolated load at zero velocity. Note that the coefficients were chosen so that the curves above gave the best fit to important average parameters (e.g. maximum power, force at a given V). Hence they differ somewhat from the average coefficients quoted by Rome and Sosnicki (1990; a hyperbolic curve with average coefficients may be skewed and may not accurately represent the average shape). comparing V and the force-velocity characteristics of red muscle should enable us to quantify the relative numbers of fibres recruited at each temperature. To swim steadily at a given speed, the carp's muscles must generate the same total power at both 10 and 20 C. Because V is independent of temperature, this means the muscle must generate the same total force (i.e. power=forcexvelocity). Fig. 7 shows that, at a swimming speed of 20cms" 1 (V=0.95MLs~ 1, where ML is muscle length), the muscle would generate 60kNm~ 2 at 10 C and 85kNm~ 2 at 20 C. Thus, the fish must recruit 1.42-fold greater fibre cross-section at 10 C to generate the same total force. This ratio increases as swimming speed increases. At 30 cm s~* (V= 1.28 ML s" x ), the red muscle at 10 C generates 46 kn irt 2 and at 20 C generates 70kNm~ 2 ; thus 1.53-fold greater fibre cross-section must be recruited at 10 C than at 20 C. Although it is clear that at 10 C the carp must recruit all its red motor units (and hence start to recruit its white ones) at a lower speed of swimming, the comparison of V and V max predicts the swimming speed at which this occurs at different temperatures. Fig. 7 shows that recruitment of white muscle occurred at both temperatures at a V where red muscle power output started to decline (right-hand ^vertical dotted lines; 1.28MLs" 1 at 10 C and 2.04MLs" 1 at 20 C) in the face of the requirement of generating additional power to swim faster. The 1.6-fold higher V at 20 C than at 10 C is matched by a Q 10 of 1.63 for V max and thus the

14 176 L. C. ROME, R. P. FUNKE AND R. MCN. ALEXANDER recruitment of white muscle occurred at both temperatures when V/V max of the red muscle reached a value of Therefore, because of its lower V max, cold carp must recruit its white muscle at slower swimming speeds. Burst-and-coast swimming at high speeds with white muscle In addition to a possible hydrodynamic advantage (Weihs, 1974; Videler and Weihs, 1982), an energetic advantage may be realized by the muscle fibres of carp during burst-and-coast swimming. Although energetics measurements were not made in this study, the efficiency of fish muscle is known to decrease from a maximal value as V/V max drops below about 0.2 (Curtin and Woledge, 1988). The carp in the present experiments appear to adopt a burst-and-coast pattern of swimming when the major muscle fibre type used would have too small a V/V max to generate power with a high efficiency if the fish were swimming steadily. The burst-and-coast pattern allows the fish to make the normal sarcomere excursion in a shorter time (shorter duty cycle), resulting in a higher V (and V/V max ). For instance, carp used the burst-and-coast mode of swimming at high swimming speeds when the white muscle was recruited. Because of its higher V max and two- to fourfold greater gear ratio (Rome et al. 1988; L. C. Rome and A. A. Sosnicki, in preparation), if the fish were to continue swimming steadily the white muscle would be shortening with a low V/V max. Thus, the fish makes rapid tail beats with white muscle and obtains a two- to threefold higher V (G. J. Lutz and L. C. Rome, unpublished finding) and presumably a more optimal V/V max. Burst-and-coast swimming at low speed with red muscle Carp also switch to burst-and-coast swimming at low swimming speeds (at both temperatures) where V/V max in the red muscle decreased below Because Vmax was 39 % lower at 10 C than at 20 C, at 10 C carp could use its red fibres to a 36 % lower V. Hence, at 10 C the carp could swim steadily at 15 cms' 1, whereas at 20 C the carp used burst-and-coast swimming at speeds below 20cms" 1, possibly to maintain a relatively higher V/V max. V/V max : a design constraint of muscle V/V max appears to set both the lowest and highest velocities over which a fibre can be effectively used during locomotion (i.e. V/V max = ) at both low and high temperatures. Previously, Rome etal. (1988) showed that, at a given temperature, slow red and fast white muscle are used over a similar range of V/V max, despite their 2.6-fold difference in V max. In any particular fibre arrangement, it is likely that the parameters that limit the range of V over which a given muscle fibre type is used are the reduced efficiency at low V/V max and the reduced power output at high V/V max. Despite the large differences in V max between fibre types and at different temperatures, muscles are used precisely over the same range of V/V max. This suggests that V/V max is an important constraint. V/V max is not the only design constraint in muscle. L. C. Rome and A. A. Sosnicki (in preparation) have shown that myofilament overlap is an important
, Marsh and Bennett (e.g. Marsh and Bennett, 1985; Marsh, 1988), and Stevens (e.g. 1988) suggests that the kinetics of activation and relaxation may be important as well.")
15 Muscle velocity in swimming carp 111 constraint; that is, fibres are anatomically arranged so that nearly maximal overlap is maintained during all movements. In addition, the extensive work of Josephson (e.g. 1985), Marsh and Bennett (e.g. Marsh and Bennett, 1985; Marsh, 1988), and Stevens (e.g. 1988) suggests that the kinetics of activation and relaxation may be important as well. By reproducing in vitro the exact stimulation pattern and length changes the muscle undergoes in vivo, we should be able to evaluate their relative importance. The authors thank Dr Marvin Freadman for use of his swimming machine in Woods Hole and Dr In-ho Choi for help with the figures. Research was supported by grants to LCR from the Whitaker Foundation, NIH (AR38404), University of Pennsylvania Research Foundation and University of Pennsylvania BRSG. References BAINBRIDGE, R. (1963). Caudal fin and body movement in the propulsion of some fish. J. exp. Biol. 40, BENNETT, A. F. (1984). Thermal dependence of muscle function. Am. J. Physiol. 247, R217-R229. CURTIN, N. A. AND WOLEDGE, R. C. (1988). Energetic cost of power output by isolated fibre bundles from dogfish white muscle. J. Physiol., Lond. 407, 74P. HUNTER, J. R. AND ZWEIFFEL, J. R. (1971). Swimming speed, tail beat frequency, tail beat amplitude, and size in jack mackerel and other fishes. Fish. Bull. 69, JAYNE, B. C, BENNETT, A. F. AND LAUDER, G. V. (1988). Effects of temperature on muscle 1 activity during lizard locomotion. Am. Zool. 28, 15A. JOSEPHSON, R. K. (1985). The mechanical power output of a tettigoniid wing muscle during singing and flight. J. exp. Biol. 117, LANCZOS, C. (1956). Applied Analysis. Englewood Cliff, NJ: Prentice Hall Inc. MARSH, R. L. (1988). Ontogenesis of contractile properties of skeletal muscle and sprint performance in the lizard Dipsosaurus dorsalis. J. exp. Biol. 137, MARSH, R. L. AND BENNETT, A. F. (1985). Thermal dependence of isotonic contractile properties of skeletal muscle and sprint performance of the lizard Dipsosaurus dorsalis. J. comp. Physiol. B 155, ROME, L. C. (1982). The energetic cost of running with different muscle temperatures in savannah monitor lizards. J. exp. Biol. 97, ROME, L. C. (1986). The influence of temperature on muscle and locomotory performance. In Living in the Cold: Physiological and Biochemical Adaptations (ed. H. C. Heller, H. J. Musacchia and L. C. H. Wang), pp New York: Elsevier. ROME, L. C. (1990). The influence of temperature on muscle recruitment and function in vivo. Am. J. Physiol. in press. ROME, L. C, FUNKE, R. P., ALEXANDER, R. MCN., LUTZ, G., ALDRIDGE, H. D. J. N., SCOTT, F. AND FREADMAN, M. (1988). Why animals have different muscle fibre types. Nature 355, ROME, L. C, LOUGHNA, P. T. ANDGOLDSPINK, G. (1984). Musclefiberrecruitment as a function of swim speed and muscle temperature in carp. Am. J. Physiol. 247, R272-R279. ROME, L. C, LOUGHNA, P. T. AND GOLDSPINK, G. (1985). Temperature acclimation improves sustained swimming performance at low temperatures in carp. Science 228, ROME, L. C. AND SOSNICKI, A. A. (1990). The influence of temperature on mechanics of red muscle in carp. J. Physiol., Lond. 427, SISSON, J. E. AND SIDELL, B. D. (1987). Effect of thermal acclimation on muscle fiber recruitment of swimming striped bass (Morone saxatilis). Physiol. Zool. 60, SMIT, H., AMELINK-KOUSTALL, J. M., VIJVERBERG, J. AND VON VAUPEL-KLEIN, J. C. (1971).
.")
16 178 L. C. ROME, R. P. FUNKE AND R. MCN. ALEXANDER Oxygen consumption and efficiency of swimming goldfish. Comp. Biochem. Physiol. 39A, STEVENS, E. D. (1979). The effect of temperature on tail beat frequency of fish swimming at constant velocity. Can. J. Zool. 57, STEVENS, E. D. (1988). Effect of ph and stimulus phase on work done by isolated frog sartorius muscle during cyclical contraction. J. Muscle Res. Cell Motil. 9, VIDELER, J. J. (1981). Swimming movements, body structure, and propulsion in cod (Gadus morhua). In Vertebrate Locomotion, Symposium of the Zoological Society of London, no. 48 (ed. M. H. Day), pp London: Academic Press. VIDELER, J. J. AND WEIHS, D. (1982). Energetic advantages of burst-and-coast swimming of fish at high speeds. J. exp. Biol. 97, WARDLE, C. S. (1975). Limit offish swimming speed. Nature 255, WEIHS, D. (1974). Energetic advantages of burst swimming offish.j. theor. Biol. 48,
SIMULTANEOUS RECORDINGS OF VELOCITY AND VIDEO DURING SWIMMING
 Portuguese Journal of Sport Sciences. 6:supl. 2, 32-35, 2006 SIMULTANEOUS RECORDINGS OF VELOCITY AND VIDEO DURING SWIMMING Albert B. Craig 1, Budd Termin2, and David R. Pendergast 2 1University of Rochester,
Portuguese Journal of Sport Sciences. 6:supl. 2, 32-35, 2006 SIMULTANEOUS RECORDINGS OF VELOCITY AND VIDEO DURING SWIMMING Albert B. Craig 1, Budd Termin2, and David R. Pendergast 2 1University of Rochester,
Characterizers for control loops
 Characterizers for control loops By: F. G. Shinskey (May 1999) Introduction Commercial controllers such as the PID series (proportional, integral, derivative, and their combinations) are linear devices
Characterizers for control loops By: F. G. Shinskey (May 1999) Introduction Commercial controllers such as the PID series (proportional, integral, derivative, and their combinations) are linear devices
The Variation of Muscle Oxygen Consumption With Velocity of Shortening
 The Variation of Muscle Oxygen Consumption With Velocity of Shortening R.J. BASKIN From the Department of Zoology, University of California, Davis ABSTRACT Total oxygen consumption following contraction
The Variation of Muscle Oxygen Consumption With Velocity of Shortening R.J. BASKIN From the Department of Zoology, University of California, Davis ABSTRACT Total oxygen consumption following contraction
Wave phenomena in a ripple tank
 Wave phenomena in a ripple tank LEP Related topics Generation of surface waves, propagation of surface waves, reflection of waves, refraction of waves, Doppler Effect. Principle Water waves are generated
Wave phenomena in a ripple tank LEP Related topics Generation of surface waves, propagation of surface waves, reflection of waves, refraction of waves, Doppler Effect. Principle Water waves are generated
ZIN Technologies PHi Engineering Support. PHi-RPT CFD Analysis of Large Bubble Mixing. June 26, 2006
 ZIN Technologies PHi Engineering Support PHi-RPT-0002 CFD Analysis of Large Bubble Mixing Proprietary ZIN Technologies, Inc. For nearly five decades, ZIN Technologies has provided integrated products and
ZIN Technologies PHi Engineering Support PHi-RPT-0002 CFD Analysis of Large Bubble Mixing Proprietary ZIN Technologies, Inc. For nearly five decades, ZIN Technologies has provided integrated products and
WHITE MUSCLE STRAIN IN THE COMMON CARP AND RED TO WHITE MUSCLE GEARING RATIOS IN FISH
 The Journal of Experimental Biology 202, 521 528 (1999) Printed in Great Britain The Company of Biologists Limited 1999 JEB1688 521 WHITE MUSCLE STRAIN IN THE COMMON CARP AND RED TO WHITE MUSCLE GEARING
The Journal of Experimental Biology 202, 521 528 (1999) Printed in Great Britain The Company of Biologists Limited 1999 JEB1688 521 WHITE MUSCLE STRAIN IN THE COMMON CARP AND RED TO WHITE MUSCLE GEARING
Atmospheric Waves James Cayer, Wesley Rondinelli, Kayla Schuster. Abstract
 Atmospheric Waves James Cayer, Wesley Rondinelli, Kayla Schuster Abstract It is important for meteorologists to have an understanding of the synoptic scale waves that propagate thorough the atmosphere
Atmospheric Waves James Cayer, Wesley Rondinelli, Kayla Schuster Abstract It is important for meteorologists to have an understanding of the synoptic scale waves that propagate thorough the atmosphere
INSTRUMENT INSTRUMENTAL ERROR (of full scale) INSTRUMENTAL RESOLUTION. Tutorial simulation. Tutorial simulation
 INSTRUMENTAL RESOLUTION. Tutorial simulation. Tutorial simulation") Lab 1 Standing Waves on a String Learning Goals: To distinguish between traveling and standing waves To recognize how the wavelength of a standing wave is measured To recognize the necessary conditions
Lab 1 Standing Waves on a String Learning Goals: To distinguish between traveling and standing waves To recognize how the wavelength of a standing wave is measured To recognize the necessary conditions
CAUDAL FIN AND BODY MOVEMENT IN THE PROPULSION OF SOME FISH
 J. Exp. Biol. (1963), 40. 23-56 23 With 1 plate and 21 text-figures Printed in Great Britain CAUDAL FIN AND BODY MOVEMENT IN THE PROPULSION OF SOME FISH BY RICHARD BAINBRIDGE Zoological Laboratory, Cambridge
J. Exp. Biol. (1963), 40. 23-56 23 With 1 plate and 21 text-figures Printed in Great Britain CAUDAL FIN AND BODY MOVEMENT IN THE PROPULSION OF SOME FISH BY RICHARD BAINBRIDGE Zoological Laboratory, Cambridge
Flow in a shock tube
 Flow in a shock tube April 30, 05 Summary In the lab the shock Mach number as well as the Mach number downstream the moving shock are determined for different pressure ratios between the high and low pressure
Flow in a shock tube April 30, 05 Summary In the lab the shock Mach number as well as the Mach number downstream the moving shock are determined for different pressure ratios between the high and low pressure
Aalborg Universitet. Published in: Proceedings of Offshore Wind 2007 Conference & Exhibition. Publication date: 2007
 Aalborg Universitet Design Loads on Platforms on Offshore wind Turbine Foundations with Respect to Vertical Wave Run-up Damsgaard, Mathilde L.; Gravesen, Helge; Andersen, Thomas Lykke Published in: Proceedings
Aalborg Universitet Design Loads on Platforms on Offshore wind Turbine Foundations with Respect to Vertical Wave Run-up Damsgaard, Mathilde L.; Gravesen, Helge; Andersen, Thomas Lykke Published in: Proceedings
IS THE HIGH COST OF BODY/CAUDAL FIN UNDULATORY SWIMMING DUE TO INCREASED FRICTION DRAG OR INERTIAL RECOIL?
 /. exp. Biol. 162, 157-166 (1992) 157 Printed in Great Britain The Company of Biologists Limited 1992 IS THE HIGH COST OF BODY/CAUDAL FIN UNDULATORY SWIMMING DUE TO INCREASED FRICTION DRAG OR INERTIAL
/. exp. Biol. 162, 157-166 (1992) 157 Printed in Great Britain The Company of Biologists Limited 1992 IS THE HIGH COST OF BODY/CAUDAL FIN UNDULATORY SWIMMING DUE TO INCREASED FRICTION DRAG OR INERTIAL
plethysmographic methods that when the subject was pinched on the upper
 24 J. Physiol. (I95I) II2, 24-2I 6I2.I5.6II.976 THE DECREASE IN HAND BLOOD FLOW FOLLOWING INFLATION OF AN ARTERIAL OCCLUSION CUFF ON THE OPPOSITE ARM BY IAN C. RODDIE From the Department of Physiology,
24 J. Physiol. (I95I) II2, 24-2I 6I2.I5.6II.976 THE DECREASE IN HAND BLOOD FLOW FOLLOWING INFLATION OF AN ARTERIAL OCCLUSION CUFF ON THE OPPOSITE ARM BY IAN C. RODDIE From the Department of Physiology,
Fish Biorobotics. Fishes as model systems for understanding aquatic propulsion. George V. Lauder Harvard University
 Fish Biorobotics Fishes as model systems for understanding aquatic propulsion 28,000 species of fishes Half of all vertebrates are fishes Fishes are 550 million years old Tremendous opportunity for selection
Fish Biorobotics Fishes as model systems for understanding aquatic propulsion 28,000 species of fishes Half of all vertebrates are fishes Fishes are 550 million years old Tremendous opportunity for selection
WAVES. Pulses are disturbances or a single wave motion. A continuous production of pulses will give rise to a progressive wave (wave train).
.") 1 WAVES Types of Waves Pulses Pulses are disturbances or a single wave motion. A continuous production of pulses will give rise to a progressive wave (wave train). Progressive Waves A progressive wave
1 WAVES Types of Waves Pulses Pulses are disturbances or a single wave motion. A continuous production of pulses will give rise to a progressive wave (wave train). Progressive Waves A progressive wave
ARE YOU A SLOW- OR A FAST-TWITCH RUNNER?
 ARE YOU A SLOW- OR A FAST-TWITCH RUNNER? How can we individualize our training towards our different goals based on our muscular makeup? In this article you will learn how to decide if you are a Fast-twitch,
ARE YOU A SLOW- OR A FAST-TWITCH RUNNER? How can we individualize our training towards our different goals based on our muscular makeup? In this article you will learn how to decide if you are a Fast-twitch,
*Present address: University of Durham, Department of Biological Sciences, South Road, Durham DH1 3LE, UK.
 J. exp. Biol. 175, 325 331 (1993) Printed in Great Britain The Company of Biologists Limited 1993 325 SHORT COMMUNICATION AN ANNULAR RESPIROMETER FOR MEASURING AEROBIC METABOLIC RATES OF LARGE, SCHOOLING
J. exp. Biol. 175, 325 331 (1993) Printed in Great Britain The Company of Biologists Limited 1993 325 SHORT COMMUNICATION AN ANNULAR RESPIROMETER FOR MEASURING AEROBIC METABOLIC RATES OF LARGE, SCHOOLING
AC : ACCURATE CRYOCHAMBER FOR A SMALL LABORATORY WITH SMALL BUDGET
 AC 2007-1108: ACCURATE CRYOCHAMBER FOR A SMALL LABORATORY WITH SMALL BUDGET Matthew Braley, University of Idaho Paul Anderson, University of Idaho Tracey Windley, University of Idaho Kevin Buck, University
AC 2007-1108: ACCURATE CRYOCHAMBER FOR A SMALL LABORATORY WITH SMALL BUDGET Matthew Braley, University of Idaho Paul Anderson, University of Idaho Tracey Windley, University of Idaho Kevin Buck, University
SEMATECH Provisional Test Method for Pressure Cycle Testing Filter Cartridges Used in UPW Distribution Systems
 Provisional Test Method for Pressure Cycle Testing Filter Cartridges Used in UPW Distribution Systems Technology Transfer 92010941B-STD and the logo are registered service marks of, Inc. 1996, Inc. Provisional
Provisional Test Method for Pressure Cycle Testing Filter Cartridges Used in UPW Distribution Systems Technology Transfer 92010941B-STD and the logo are registered service marks of, Inc. 1996, Inc. Provisional
EXPERIMENTAL STUDY ON THE DISCHARGE CHARACTERISTICS OF SLUICE FOR TIDAL POWER PLANT
 EXPERIMENTAL STUDY ON THE DISCHARGE CHARACTERISTICS OF SLUICE FOR TIDAL POWER PLANT Sang-Ho Oh 1, Kwang Soo Lee 1 and Dal Soo Lee 1 The discharge characteristics of sluice caisson for tidal power plant
EXPERIMENTAL STUDY ON THE DISCHARGE CHARACTERISTICS OF SLUICE FOR TIDAL POWER PLANT Sang-Ho Oh 1, Kwang Soo Lee 1 and Dal Soo Lee 1 The discharge characteristics of sluice caisson for tidal power plant
Swimming Stroke Mechanics
 Swimming Stroke Mechanics What we continue to learn from High-speed Videography and Biomechanical Motion Analysis Jan Prins, Ph.D. Aquatic Research Laboratory University of Hawaii Swimming Biomechanics,
Swimming Stroke Mechanics What we continue to learn from High-speed Videography and Biomechanical Motion Analysis Jan Prins, Ph.D. Aquatic Research Laboratory University of Hawaii Swimming Biomechanics,
Physiological Assessment: Summary Report 11 December 2011
 Physiological Assessment: Summary Report 11 December 211 Katie Parry MD/LD Runner Athlete Test conducted and report compiled by: Dr Andi Drake England Athletics, Leeds Metropolitan University Headingley
Physiological Assessment: Summary Report 11 December 211 Katie Parry MD/LD Runner Athlete Test conducted and report compiled by: Dr Andi Drake England Athletics, Leeds Metropolitan University Headingley
Micro Motion Pressure Drop Testing
 12/2018 Micro Motion Pressure Drop Testing www.emerson.com/micromotion Introduction Micro Motion has traditionally taken a very conservative approach to pressure drop, with single pressure measurements
12/2018 Micro Motion Pressure Drop Testing www.emerson.com/micromotion Introduction Micro Motion has traditionally taken a very conservative approach to pressure drop, with single pressure measurements
Exploring the relationship between the pressure of the ball and coefficient of restitution.
 Exploring the relationship between the pressure of the ball and coefficient of restitution. When I started thinking about possible investigations I knew I wanted to create a lab that was related to sports.
Exploring the relationship between the pressure of the ball and coefficient of restitution. When I started thinking about possible investigations I knew I wanted to create a lab that was related to sports.
Hydrodynamic function of dorsal and anal fins in brook trout (Salvelinus fontinalis)
") Hydrodynamic function of dorsal and anal fins in brook trout (Salvelinus fontinalis) The Harvard community has made this article openly available. Please share how this access benefits you. Your story
Hydrodynamic function of dorsal and anal fins in brook trout (Salvelinus fontinalis) The Harvard community has made this article openly available. Please share how this access benefits you. Your story
Movement and Position
 Movement and Position Syllabus points: 1.2 plot and interpret distance-time graphs 1.3 know and use the relationship between average speed, distance moved and 1.4 describe experiments to investigate the
Movement and Position Syllabus points: 1.2 plot and interpret distance-time graphs 1.3 know and use the relationship between average speed, distance moved and 1.4 describe experiments to investigate the
APPROACH RUN VELOCITIES OF FEMALE POLE VAULTERS
 APPROACH RUN VELOCITIES OF FEMALE POLE VAULTERS Peter M. McGinnis, Physical Education Department, SUNY College at Cortland, Cortland, New York INTRODUCTION Running speed is an important determinant of
APPROACH RUN VELOCITIES OF FEMALE POLE VAULTERS Peter M. McGinnis, Physical Education Department, SUNY College at Cortland, Cortland, New York INTRODUCTION Running speed is an important determinant of
Analysis of Pressure Rise During Internal Arc Faults in Switchgear
 Analysis of Pressure Rise During Internal Arc Faults in Switchgear ASANUMA, Gaku ONCHI, Toshiyuki TOYAMA, Kentaro ABSTRACT Switchgear include devices that play an important role in operations such as electric
Analysis of Pressure Rise During Internal Arc Faults in Switchgear ASANUMA, Gaku ONCHI, Toshiyuki TOYAMA, Kentaro ABSTRACT Switchgear include devices that play an important role in operations such as electric
Describing a journey made by an object is very boring if you just use words. As with much of science, graphs are more revealing.
 Distance vs. Time Describing a journey made by an object is very boring if you just use words. As with much of science, graphs are more revealing. Plotting distance against time can tell you a lot about
Distance vs. Time Describing a journey made by an object is very boring if you just use words. As with much of science, graphs are more revealing. Plotting distance against time can tell you a lot about
60bbm and less 60-50bbm 50-30bbm 30-20bbm 20-10bbm (incorporates a significant aerobic component)
") TRAINING SPECIFICS IN RELATION TO AEROBIC & ANAEROBIC CONDITIONING OF SWIMMERS By Leigh Nugent, National Youth Coach Training specificity is a term mentioned often in the literature published on the physiological
TRAINING SPECIFICS IN RELATION TO AEROBIC & ANAEROBIC CONDITIONING OF SWIMMERS By Leigh Nugent, National Youth Coach Training specificity is a term mentioned often in the literature published on the physiological
OXYGEN POWER By Jack Daniels, Jimmy Gilbert, 1979
 1 de 6 OXYGEN POWER By Jack Daniels, Jimmy Gilbert, 1979 Appendix A Running is primarily a conditioning sport. It is not a skill sport as is basketball or handball or, to a certain extent, swimming where
1 de 6 OXYGEN POWER By Jack Daniels, Jimmy Gilbert, 1979 Appendix A Running is primarily a conditioning sport. It is not a skill sport as is basketball or handball or, to a certain extent, swimming where
Sizing Pulsation Dampeners Is Critical to Effectiveness
 Sizing Pulsation Dampeners Is Critical to Effectiveness Pressure variation is an important consideration when determining the appropriate size pulsation dampener needed for an application. by David McComb,
Sizing Pulsation Dampeners Is Critical to Effectiveness Pressure variation is an important consideration when determining the appropriate size pulsation dampener needed for an application. by David McComb,
Cover Page for Lab Report Group Portion. Pump Performance
 Cover Page for Lab Report Group Portion Pump Performance Prepared by Professor J. M. Cimbala, Penn State University Latest revision: 02 March 2012 Name 1: Name 2: Name 3: [Name 4: ] Date: Section number:
Cover Page for Lab Report Group Portion Pump Performance Prepared by Professor J. M. Cimbala, Penn State University Latest revision: 02 March 2012 Name 1: Name 2: Name 3: [Name 4: ] Date: Section number:
LOW PRESSURE EFFUSION OF GASES revised by Igor Bolotin 03/05/12
 LOW PRESSURE EFFUSION OF GASES revised by Igor Bolotin 03/05/ This experiment will introduce you to the kinetic properties of low-pressure gases. You will make observations on the rates with which selected
LOW PRESSURE EFFUSION OF GASES revised by Igor Bolotin 03/05/ This experiment will introduce you to the kinetic properties of low-pressure gases. You will make observations on the rates with which selected
UNIT 15 WATER HAMMER AND SURGE TANKS
 UNT 15 WATER HAMMER AND SURGE TANKS Structure 15.1 ntroduction Objectives 15.2 Water Hammer 15.2.1 Expression for Rise in Pressure 15.3 Rapid Acceleration of Flow 15.4 Surge Tanks 15.5 Summary 15.6 Keywords
UNT 15 WATER HAMMER AND SURGE TANKS Structure 15.1 ntroduction Objectives 15.2 Water Hammer 15.2.1 Expression for Rise in Pressure 15.3 Rapid Acceleration of Flow 15.4 Surge Tanks 15.5 Summary 15.6 Keywords
Sprint Hurdles SPRINT HURDLE RACES: A KINEMATIC ANALYSIS
 Sprint Hurdles SPRINT HURDLE RACES: A KINEMATIC ANALYSIS By Pierre-Jean Vazel, December 2011 In order to prescribe the most effective training possible it is important for coaches to have a sound understanding
Sprint Hurdles SPRINT HURDLE RACES: A KINEMATIC ANALYSIS By Pierre-Jean Vazel, December 2011 In order to prescribe the most effective training possible it is important for coaches to have a sound understanding
485 Annubar Primary Flow Element Installation Effects
 ROSEMOUNT 485 ANNUBAR 485 Annubar Primary Flow Element Installation Effects CONTENTS Mounting hole diameter Alignment error Piping Geometry Induced Flow Disturbances Pipe reducers and expansions Control
ROSEMOUNT 485 ANNUBAR 485 Annubar Primary Flow Element Installation Effects CONTENTS Mounting hole diameter Alignment error Piping Geometry Induced Flow Disturbances Pipe reducers and expansions Control
The Incremental Evolution of Gaits for Hexapod Robots
 The Incremental Evolution of Gaits for Hexapod Robots Abstract Gait control programs for hexapod robots are learned by incremental evolution. The first increment is used to learn the activations required
The Incremental Evolution of Gaits for Hexapod Robots Abstract Gait control programs for hexapod robots are learned by incremental evolution. The first increment is used to learn the activations required
AN ANALYSIS ON HIGH PRESSURE DYNAMIC CALIBRATORS USED IN THE DEFENSE AREAS
 AN ANALYSIS ON HIGH PRESSURE DYNAMIC CALIBRATORS USED IN THE DEFENSE AREAS Sung Soo HongPresenter Agency for Defense Development, Taean, 357-942, South Korea sungsoo@add.re.kr Abstract Up to now, there
AN ANALYSIS ON HIGH PRESSURE DYNAMIC CALIBRATORS USED IN THE DEFENSE AREAS Sung Soo HongPresenter Agency for Defense Development, Taean, 357-942, South Korea sungsoo@add.re.kr Abstract Up to now, there
Define transverse waves and longitudinal waves. Draw a simple diagram of each
 AP Physics Study Guide Chapters 11, 12, 24 Waves, Sound, Light & Interference Name Write the equation that defines each quantity, include units for all quantities. wave speed-wavelength equation natural
AP Physics Study Guide Chapters 11, 12, 24 Waves, Sound, Light & Interference Name Write the equation that defines each quantity, include units for all quantities. wave speed-wavelength equation natural
Components of the 400m Hurdles
 Components of the 400m Hurdles By Ray Boyd NATIONAL 400M HURDLE COACH-AUSTRALIA FROM: Modern Athlete and Coach The 400m hurdles race can be broken up into four stages: Start to First Hurdle Movement Across
Components of the 400m Hurdles By Ray Boyd NATIONAL 400M HURDLE COACH-AUSTRALIA FROM: Modern Athlete and Coach The 400m hurdles race can be broken up into four stages: Start to First Hurdle Movement Across
Evaluation of aerodynamic criteria in the design of a small wind turbine with the lifting line model
 Evaluation of aerodynamic criteria in the design of a small wind turbine with the lifting line model Nicolas BRUMIOUL Abstract This thesis deals with the optimization of the aerodynamic design of a small
Evaluation of aerodynamic criteria in the design of a small wind turbine with the lifting line model Nicolas BRUMIOUL Abstract This thesis deals with the optimization of the aerodynamic design of a small
Experiment 8: Minor Losses
 Experiment 8: Minor Losses Purpose: To determine the loss factors for flow through a range of pipe fittings including bends, a contraction, an enlargement and a gate-valve. Introduction: Energy losses
Experiment 8: Minor Losses Purpose: To determine the loss factors for flow through a range of pipe fittings including bends, a contraction, an enlargement and a gate-valve. Introduction: Energy losses
Ripple Tank Exploring the Properties of Waves Using a Ripple Tank
 Exploring the Properties of Waves Using a The ripple tank is a shallow, glass-bottomed container that is filled with water to a depth of 1 or 2 centimeters. There is a light source that is placed above
Exploring the Properties of Waves Using a The ripple tank is a shallow, glass-bottomed container that is filled with water to a depth of 1 or 2 centimeters. There is a light source that is placed above
Conventional Ship Testing
 Conventional Ship Testing Experimental Methods in Marine Hydrodynamics Lecture in week 34 Chapter 6 in the lecture notes 1 Conventional Ship Testing - Topics: Resistance tests Propeller open water tests
Conventional Ship Testing Experimental Methods in Marine Hydrodynamics Lecture in week 34 Chapter 6 in the lecture notes 1 Conventional Ship Testing - Topics: Resistance tests Propeller open water tests
LocoMorph Deliverable 4.4 part 2.4
 LocoMorph Deliverable 4.4 part 2.4 Kinematics in healthy and morphosed long-tailed lizards (Takydromus sexlineatus): comparison of a simulation model with experimental animal data. Aim D Août K, Karakasiliotis
LocoMorph Deliverable 4.4 part 2.4 Kinematics in healthy and morphosed long-tailed lizards (Takydromus sexlineatus): comparison of a simulation model with experimental animal data. Aim D Août K, Karakasiliotis
The Aerodynamic Drag of Parafoils
 The Aerodynamic Drag of Parafoils A. C. Carruthers and A. Filippone The University of Manchester Manchester M60 1QD United Kingdom Introduction The parafoil is an aerodynamic decelerator that uses the
The Aerodynamic Drag of Parafoils A. C. Carruthers and A. Filippone The University of Manchester Manchester M60 1QD United Kingdom Introduction The parafoil is an aerodynamic decelerator that uses the
46 Chapter 8 Statistics: An Introduction
 46 Chapter 8 Statistics: An Introduction Activity 5 Continued Box 4 1. The median is 6. The mode is 5. The mean is about 7 (6.8). 2. The median and the mode were unchanged, but the mean increased significantly.
46 Chapter 8 Statistics: An Introduction Activity 5 Continued Box 4 1. The median is 6. The mode is 5. The mean is about 7 (6.8). 2. The median and the mode were unchanged, but the mean increased significantly.
CHEMICAL ENGINEERING LABORATORY CHEG 239W. Control of a Steam-Heated Mixing Tank with a Pneumatic Process Controller
 CHEMICAL ENGINEERING LABORATORY CHEG 239W Control of a Steam-Heated Mixing Tank with a Pneumatic Process Controller Objective The experiment involves tuning a commercial process controller for temperature
CHEMICAL ENGINEERING LABORATORY CHEG 239W Control of a Steam-Heated Mixing Tank with a Pneumatic Process Controller Objective The experiment involves tuning a commercial process controller for temperature
Functional Morphology of Locomotion and Feeding. Functional Morphology of Locomotion and Feeding
 Functional Morphology of Structure and Function tied together Locomotion: Terrestrial = gravity Aquatic = density of water and drag exerted by it remember 800xdense and 50x viscous. Energetically it is
Functional Morphology of Structure and Function tied together Locomotion: Terrestrial = gravity Aquatic = density of water and drag exerted by it remember 800xdense and 50x viscous. Energetically it is
Dynamics of bubble rising at small Reynolds numbers
 MATEC Web of Conferences 3, 01004 ( 015) DOI: 10.1051/ matecconf/ 015301004 C Owned by the authors, published by EDP Sciences, 015 Dynamics of bubble rising at small Reynolds numbers Vladimir Arkhipov
MATEC Web of Conferences 3, 01004 ( 015) DOI: 10.1051/ matecconf/ 015301004 C Owned by the authors, published by EDP Sciences, 015 Dynamics of bubble rising at small Reynolds numbers Vladimir Arkhipov
EFFICIENCY OF UPHILL LOCOMOTION IN NOCTURNAL AND DIURNAL LIZARDS
 The Journal of Experimental Biology 199, 587 592 (1996) Printed in Great Britain The Company of Biologists Limited 1996 JEB0066 587 EFFICIENCY OF UPHILL LOCOMOTION IN NOCTURNAL AND DIURNAL LIZARDS C. T.
The Journal of Experimental Biology 199, 587 592 (1996) Printed in Great Britain The Company of Biologists Limited 1996 JEB0066 587 EFFICIENCY OF UPHILL LOCOMOTION IN NOCTURNAL AND DIURNAL LIZARDS C. T.
Technical Note. Determining the surface tension of liquids by measurements on pendant drops
 Technical Note Pendant Drop Measurements Technical note: TN316e Industry section: all Author: FT, TW Date: 12/2010 Method: Drop Shape Analyzer DSA100 Keywords: Methods, surface tension, interfacial tension,
Technical Note Pendant Drop Measurements Technical note: TN316e Industry section: all Author: FT, TW Date: 12/2010 Method: Drop Shape Analyzer DSA100 Keywords: Methods, surface tension, interfacial tension,
SUBMERGED VENTURI FLUME. Tom Gill 1 Robert Einhellig 2 ABSTRACT
 SUBMERGED VENTURI FLUME Tom Gill 1 Robert Einhellig 2 ABSTRACT Improvement in canal operating efficiency begins with establishing the ability to measure flow at key points in the delivery system. The lack
SUBMERGED VENTURI FLUME Tom Gill 1 Robert Einhellig 2 ABSTRACT Improvement in canal operating efficiency begins with establishing the ability to measure flow at key points in the delivery system. The lack
HOW FISH POWER PREDATION FAST-STARTS
 The Journal of Experimental Biology 198, 1851 1861 (1995) Printed in Great Britain The Company of Biologists Limited 1995 1851 HOW FISH POWER PREDATION FAST-STARTS IAN A. JOHNSTON 1, JOHAN L. VAN LEEUWEN
The Journal of Experimental Biology 198, 1851 1861 (1995) Printed in Great Britain The Company of Biologists Limited 1995 1851 HOW FISH POWER PREDATION FAST-STARTS IAN A. JOHNSTON 1, JOHAN L. VAN LEEUWEN
Waves. harmonic wave wave equation one dimensional wave equation principle of wave fronts plane waves law of reflection
 Waves Vocabulary mechanical wave pulse continuous periodic wave amplitude wavelength period frequency wave velocity phase transverse wave longitudinal wave intensity displacement wave number phase velocity
Waves Vocabulary mechanical wave pulse continuous periodic wave amplitude wavelength period frequency wave velocity phase transverse wave longitudinal wave intensity displacement wave number phase velocity
ScienceDirect. Relating baseball seam height to carry distance
 Available online at www.sciencedirect.com ScienceDirect Procedia Engineering 112 (2015 ) 406 411 7th Asia-Pacific Congress on Sports Technology, APCST 2015 Relating baseball seam height to carry distance
Available online at www.sciencedirect.com ScienceDirect Procedia Engineering 112 (2015 ) 406 411 7th Asia-Pacific Congress on Sports Technology, APCST 2015 Relating baseball seam height to carry distance
CHAPTER IV FINITE ELEMENT ANALYSIS OF THE KNEE JOINT WITHOUT A MEDICAL IMPLANT
 39 CHAPTER IV FINITE ELEMENT ANALYSIS OF THE KNEE JOINT WITHOUT A MEDICAL IMPLANT 4.1 Modeling in Biomechanics The human body, apart of all its other functions is a mechanical mechanism and a structure,
39 CHAPTER IV FINITE ELEMENT ANALYSIS OF THE KNEE JOINT WITHOUT A MEDICAL IMPLANT 4.1 Modeling in Biomechanics The human body, apart of all its other functions is a mechanical mechanism and a structure,
Lab 4: Transpiration
 Lab 4: Transpiration Water is transported in plants, from the roots to the leaves, following a decreasing water potential gradient. Transpiration, or loss of water from the leaves, helps to create a lower
Lab 4: Transpiration Water is transported in plants, from the roots to the leaves, following a decreasing water potential gradient. Transpiration, or loss of water from the leaves, helps to create a lower
The University of Hong Kong Department of Physics Experimental Physics Laboratory
 The University of Hong Kong Department of Physics Experimental Physics Laboratory PHYS2260 Heat and Waves 2260-1 LABORATORY MANUAL Experiment 1: Adiabatic Gas Law Part A. Ideal Gas Law Equipment Required:
The University of Hong Kong Department of Physics Experimental Physics Laboratory PHYS2260 Heat and Waves 2260-1 LABORATORY MANUAL Experiment 1: Adiabatic Gas Law Part A. Ideal Gas Law Equipment Required:
NUMERICAL INVESTIGATION OF THE FLOW BEHAVIOUR IN A MODERN TRAFFIC TUNNEL IN CASE OF FIRE INCIDENT
 - 277 - NUMERICAL INVESTIGATION OF THE FLOW BEHAVIOUR IN A MODERN TRAFFIC TUNNEL IN CASE OF FIRE INCIDENT Iseler J., Heiser W. EAS GmbH, Karlsruhe, Germany ABSTRACT A numerical study of the flow behaviour
- 277 - NUMERICAL INVESTIGATION OF THE FLOW BEHAVIOUR IN A MODERN TRAFFIC TUNNEL IN CASE OF FIRE INCIDENT Iseler J., Heiser W. EAS GmbH, Karlsruhe, Germany ABSTRACT A numerical study of the flow behaviour
Experimental Investigation on Changes of Water Surface Profile with Gaussian Shaped Bottom and Side Roughness
 Experimental Investigation on Changes of Water Surface Profile with Gaussian Shaped Bottom and Side Md. Rafiue Islam a, Shariful Islam b*, Md. Abdul Qaiyum Talukder c, S. M. Rezwan Hossain d Abstract Bed
Experimental Investigation on Changes of Water Surface Profile with Gaussian Shaped Bottom and Side Md. Rafiue Islam a, Shariful Islam b*, Md. Abdul Qaiyum Talukder c, S. M. Rezwan Hossain d Abstract Bed
PUBLISHED PROJECT REPORT PPR850. Optimisation of water flow depth for SCRIM. S Brittain, P Sanders and H Viner
 PUBLISHED PROJECT REPORT PPR850 Optimisation of water flow depth for SCRIM S Brittain, P Sanders and H Viner Report details Report prepared for: Project/customer reference: Copyright: Highways England,
PUBLISHED PROJECT REPORT PPR850 Optimisation of water flow depth for SCRIM S Brittain, P Sanders and H Viner Report details Report prepared for: Project/customer reference: Copyright: Highways England,
SPEED, ACCELERATION AND MANOEUVRABILITY OF TWO TELEOST FISHES
 J. exp. Biol. 102, 115122, 1983 Printed in Great Britain Company of Biologists Limited 1983 SPEED, ACCELERATION AND MANOEUVRABILITY OF TWO TELEOST FISHES BY P. W. WEBB School of Natural Resources, The
J. exp. Biol. 102, 115122, 1983 Printed in Great Britain Company of Biologists Limited 1983 SPEED, ACCELERATION AND MANOEUVRABILITY OF TWO TELEOST FISHES BY P. W. WEBB School of Natural Resources, The
Heat Engine. Reading: Appropriate sections for first, second law of thermodynamics, and PV diagrams.
 Heat Engine Equipment: Capstone, 2 large glass beakers (one for ice water, the other for boiling water), temperature sensor, pressure sensor, rotary motion sensor, meter stick, calipers, set of weights,
Heat Engine Equipment: Capstone, 2 large glass beakers (one for ice water, the other for boiling water), temperature sensor, pressure sensor, rotary motion sensor, meter stick, calipers, set of weights,
6. An oscillator makes four vibrations in one second. What is its period and frequency?
 Period and Frequency 19.1 The period of a pendulum is the time it takes to move through one cycle. As the ball on the string is pulled to one side and then let go, the ball moves to the side opposite the
Period and Frequency 19.1 The period of a pendulum is the time it takes to move through one cycle. As the ball on the string is pulled to one side and then let go, the ball moves to the side opposite the
CTB3365x Introduction to Water Treatment
 CTB3365x Introduction to Water Treatment D4b Aeration Doris van Halem Did you know that there are not just gasses in your glass of sparkling coke, but also in the tap water you drink? Welcome to the water
CTB3365x Introduction to Water Treatment D4b Aeration Doris van Halem Did you know that there are not just gasses in your glass of sparkling coke, but also in the tap water you drink? Welcome to the water
NOTES ON WATER HAMMER. 55
 NOTES ON WATER HAMMER. 55 NOTES ON WATER HAMMER. By A. B. Robison. When the flow conditions of a liquid in a pipe line are varied by the opening or closing of a valve or the equivalent, a change in the
NOTES ON WATER HAMMER. 55 NOTES ON WATER HAMMER. By A. B. Robison. When the flow conditions of a liquid in a pipe line are varied by the opening or closing of a valve or the equivalent, a change in the
Report for Experiment #11 Testing Newton s Second Law On the Moon
 Report for Experiment #11 Testing Newton s Second Law On the Moon Neil Armstrong Lab partner: Buzz Aldrin TA: Michael Collins July 20th, 1969 Abstract In this experiment, we tested Newton s second law
Report for Experiment #11 Testing Newton s Second Law On the Moon Neil Armstrong Lab partner: Buzz Aldrin TA: Michael Collins July 20th, 1969 Abstract In this experiment, we tested Newton s second law
that, as a means of progression, walking is suitable for lower speeds
 2 6I2 744.22 ENERGY EXPENDITURE IN WALKING AND RUNNING. BY M. OGASAWARA. (From the Department of Industrial Physiology, London School of Hygiene and Tropical Medicine.) (Received February 28, 1934.) IT
2 6I2 744.22 ENERGY EXPENDITURE IN WALKING AND RUNNING. BY M. OGASAWARA. (From the Department of Industrial Physiology, London School of Hygiene and Tropical Medicine.) (Received February 28, 1934.) IT
Exploring the relationship between the pressure of the ball and coefficient of restitution.
 Exploring the relationship between the pressure of the ball and coefficient of restitution. When I started thinking about possible investigations I knew I wanted to create a lab that was related to sports.
Exploring the relationship between the pressure of the ball and coefficient of restitution. When I started thinking about possible investigations I knew I wanted to create a lab that was related to sports.
9.2 Waves at Media Boundaries
 media boundary the location where two or more media meet Figure 1 The walls and shapes of recording studios are carefully designed to ensure that the sound going to the microphone is a true representation
media boundary the location where two or more media meet Figure 1 The walls and shapes of recording studios are carefully designed to ensure that the sound going to the microphone is a true representation
Autodesk Moldflow Communicator Process settings
 Autodesk Moldflow Communicator 212 Process settings Revision 1, 3 March 211. Contents Chapter 1 Process settings....................................... 1 Profiles.................................................
Autodesk Moldflow Communicator 212 Process settings Revision 1, 3 March 211. Contents Chapter 1 Process settings....................................... 1 Profiles.................................................
The Aging Curve(s) Jerry Meyer, Central Maryland YMCA Masters (CMYM)
 Jerry Meyer, Central Maryland YMCA Masters (CMYM)") The Aging Curve(s) Jerry Meyer, Central Maryland YMCA Masters (CMYM) Even the well-publicized benefits of Masters Swimming cannot prevent us from eventually slowing down as we get older. While some find
The Aging Curve(s) Jerry Meyer, Central Maryland YMCA Masters (CMYM) Even the well-publicized benefits of Masters Swimming cannot prevent us from eventually slowing down as we get older. While some find
CHAPTER 5: VACUUM TEST WITH VERTICAL DRAINS
 CHAPTER 5: VACUUM TEST WITH VERTICAL DRAINS 5.1 Introduction Using surcharging as the sole soil consolidation mean can take a long time to reach the desired soil settlement. Soil consolidation using prefabricated
CHAPTER 5: VACUUM TEST WITH VERTICAL DRAINS 5.1 Introduction Using surcharging as the sole soil consolidation mean can take a long time to reach the desired soil settlement. Soil consolidation using prefabricated
Energy balance of the model as described in Theory and Model
 Energy balance of the model as described in Theory and Model 1. Introduction 2. Power sources and -sinks 3. Numerical results 4. Final remarks 5. Heavy shell (added) 1. Introduction The energy balance
Energy balance of the model as described in Theory and Model 1. Introduction 2. Power sources and -sinks 3. Numerical results 4. Final remarks 5. Heavy shell (added) 1. Introduction The energy balance
PMT. Smaller species of annelid do not have gills. Explain why these small worms do not need gills to obtain sufficient oxygen
 1. There are many different species of annelid worm. Some are very small, only a few millimetres in length. Others, such as lugworms, are much larger. The drawing shows a lugworm and part of one of its
1. There are many different species of annelid worm. Some are very small, only a few millimetres in length. Others, such as lugworms, are much larger. The drawing shows a lugworm and part of one of its
Preliminary design of a high-altitude kite. A flexible membrane kite section at various wind speeds
 Preliminary design of a high-altitude kite A flexible membrane kite section at various wind speeds This is the third paper in a series that began with one titled A flexible membrane kite section at high
Preliminary design of a high-altitude kite A flexible membrane kite section at various wind speeds This is the third paper in a series that began with one titled A flexible membrane kite section at high
LOW PRESSURE EFFUSION OF GASES adapted by Luke Hanley and Mike Trenary
 ADH 1/7/014 LOW PRESSURE EFFUSION OF GASES adapted by Luke Hanley and Mike Trenary This experiment will introduce you to the kinetic properties of low-pressure gases. You will make observations on the
ADH 1/7/014 LOW PRESSURE EFFUSION OF GASES adapted by Luke Hanley and Mike Trenary This experiment will introduce you to the kinetic properties of low-pressure gases. You will make observations on the
Real-time Analysis of Industrial Gases with Online Near- Infrared Spectroscopy
 Real-time Analysis of Industrial Gases with Online Near- Infrared Branch 5 Petrochemical, Biofuels Keywords Process, NIR, Gases, Gas Cell, Ethylene, Carbon Dioxide, Propane, Butane, Acetylene Summary Near-infrared
Real-time Analysis of Industrial Gases with Online Near- Infrared Branch 5 Petrochemical, Biofuels Keywords Process, NIR, Gases, Gas Cell, Ethylene, Carbon Dioxide, Propane, Butane, Acetylene Summary Near-infrared
Paper 2.2. Operation of Ultrasonic Flow Meters at Conditions Different Than Their Calibration
 Paper 2.2 Operation of Ultrasonic Flow Meters at Conditions Different Than Their Calibration Mr William Freund, Daniel Measurement and Control Mr Klaus Zanker, Daniel Measurement and Control Mr Dale Goodson,
Paper 2.2 Operation of Ultrasonic Flow Meters at Conditions Different Than Their Calibration Mr William Freund, Daniel Measurement and Control Mr Klaus Zanker, Daniel Measurement and Control Mr Dale Goodson,
Time Pressure Dispensing
 Time Pressure Dispensing by Doug Dixon, GDM Product Manager What is time pressure dispensing? Time pressure is a method of dispensing liquid materials (surface mount adhesives and gasketing materials)
Time Pressure Dispensing by Doug Dixon, GDM Product Manager What is time pressure dispensing? Time pressure is a method of dispensing liquid materials (surface mount adhesives and gasketing materials)
SECTION 2 HYDROLOGY AND FLOW REGIMES
 SECTION 2 HYDROLOGY AND FLOW REGIMES In this section historical streamflow data from permanent USGS gaging stations will be presented and discussed to document long-term flow regime trends within the Cache-Bayou
SECTION 2 HYDROLOGY AND FLOW REGIMES In this section historical streamflow data from permanent USGS gaging stations will be presented and discussed to document long-term flow regime trends within the Cache-Bayou
SOLAR 93 THE 1993 AMERICAN SOLAR ENERGY SOCIETY ANNUAL CONFERENCE. Washington, DC April 22028,1993. Editors: S. M. Burley M. E.
 SOLAR 93 THE 1993 AMERICAN SOLAR ENERGY SOCIETY ANNUAL CONFERENCE Washington, DC April 22028,1993 Editors: S. M. Burley M. E. Arden American Solar Energy Society U.S. Section of the International Solar
SOLAR 93 THE 1993 AMERICAN SOLAR ENERGY SOCIETY ANNUAL CONFERENCE Washington, DC April 22028,1993 Editors: S. M. Burley M. E. Arden American Solar Energy Society U.S. Section of the International Solar
Implementing Provisions for Art. 411 of the ICR Ski Jumping
 JUMPING HILLS CONSTRUCTION NORM 2018 Implementing Provisions for Art. 411 of the ICR Ski Jumping Author: Hans-Heini Gasser (SUI) EDITION NOVEMBER 2018 Table of Contents Page 1. Preliminary Remarks 3 2.
JUMPING HILLS CONSTRUCTION NORM 2018 Implementing Provisions for Art. 411 of the ICR Ski Jumping Author: Hans-Heini Gasser (SUI) EDITION NOVEMBER 2018 Table of Contents Page 1. Preliminary Remarks 3 2.
-Elastic strain energy (duty factor decreases at higher speeds). Higher forces act on feet. More tendon stretch. More energy stored in tendon.
. Higher forces act on feet. More tendon stretch. More energy stored in tendon.") As velocity increases ( ) (i.e. increasing Froude number v 2 / gl) the component of the energy cost of transport associated with: -Internal kinetic energy (limbs accelerated to higher angular velocity).
As velocity increases ( ) (i.e. increasing Froude number v 2 / gl) the component of the energy cost of transport associated with: -Internal kinetic energy (limbs accelerated to higher angular velocity).
Gas Laws. Introduction
 Gas Laws Introduction In 1662 Robert Boyle found that, at constant temperature, the pressure of a gas and its volume are inversely proportional such that P x V = constant. This relationship is known as
Gas Laws Introduction In 1662 Robert Boyle found that, at constant temperature, the pressure of a gas and its volume are inversely proportional such that P x V = constant. This relationship is known as
BLOOD PRESSURE SENSOR BT17i USER S GUIDE
 BLOOD PRESSURE SENSOR BT17i USER S GUIDE CENTRE FOR MICROCOMPUTER APPLICATIONS http://www.cma-science.nl Short description The Blood Pressure sensor BT17i allows measuring arterial blood pressure. The
BLOOD PRESSURE SENSOR BT17i USER S GUIDE CENTRE FOR MICROCOMPUTER APPLICATIONS http://www.cma-science.nl Short description The Blood Pressure sensor BT17i allows measuring arterial blood pressure. The
Numerical and Experimental Investigation of the Possibility of Forming the Wake Flow of Large Ships by Using the Vortex Generators
 Second International Symposium on Marine Propulsors smp 11, Hamburg, Germany, June 2011 Numerical and Experimental Investigation of the Possibility of Forming the Wake Flow of Large Ships by Using the
Second International Symposium on Marine Propulsors smp 11, Hamburg, Germany, June 2011 Numerical and Experimental Investigation of the Possibility of Forming the Wake Flow of Large Ships by Using the
DETRMINATION OF A PLUNGER TYPE WAVE MAKER CHARACTERISTICE IN A TOWING TANK
 The 9 th International Conference on Coasts, Ports and Marine Structures (ICOPMAS 2010) 29 Nov.-1 Dec. 2010 (Tehran) DETRMINATION OF A PLUNGER TYPE WAVE MAKER CHARACTERISTICE IN A TOWING TANK sayed mohammad
The 9 th International Conference on Coasts, Ports and Marine Structures (ICOPMAS 2010) 29 Nov.-1 Dec. 2010 (Tehran) DETRMINATION OF A PLUNGER TYPE WAVE MAKER CHARACTERISTICE IN A TOWING TANK sayed mohammad
STUDIES IN ANIMAL LOCOMOTION
 39 1 STUDIES IN ANIMAL LOCOMOTION III. THE PROPULSIVE MECHANISM OF THE WHITING (GADUS MERLANGUS) BY J. GRAY. (From the Laboratory of Experimental Zoology, Cambridge.) {Received ^th February, 1933.) (With
39 1 STUDIES IN ANIMAL LOCOMOTION III. THE PROPULSIVE MECHANISM OF THE WHITING (GADUS MERLANGUS) BY J. GRAY. (From the Laboratory of Experimental Zoology, Cambridge.) {Received ^th February, 1933.) (With
Gerald D. Anderson. Education Technical Specialist
 Gerald D. Anderson Education Technical Specialist The factors which influence selection of equipment for a liquid level control loop interact significantly. Analyses of these factors and their interactions
Gerald D. Anderson Education Technical Specialist The factors which influence selection of equipment for a liquid level control loop interact significantly. Analyses of these factors and their interactions
1. A tendency to roll or heel when turning (a known and typically constant disturbance) 2. Motion induced by surface waves of certain frequencies.
 2. Motion induced by surface waves of certain frequencies.") Department of Mechanical Engineering Massachusetts Institute of Technology 2.14 Analysis and Design of Feedback Control Systems Fall 2004 October 21, 2004 Case Study on Ship Roll Control Problem Statement:
Department of Mechanical Engineering Massachusetts Institute of Technology 2.14 Analysis and Design of Feedback Control Systems Fall 2004 October 21, 2004 Case Study on Ship Roll Control Problem Statement:
CubeSat Balloon Drag Devices: Meeting the 25-Year De-Orbit Requirement
 CubeSat Balloon Drag Devices: Meeting the 25-Year De-Orbit Requirement Jerry K. Fuller, David Hinkley, and Siegfried W. Janson The Aerospace Corporation Physical Science Laboratories August, 2010 The Aerospace
CubeSat Balloon Drag Devices: Meeting the 25-Year De-Orbit Requirement Jerry K. Fuller, David Hinkley, and Siegfried W. Janson The Aerospace Corporation Physical Science Laboratories August, 2010 The Aerospace
PIG MOTION AND DYNAMICS IN COMPLEX GAS NETWORKS. Dr Aidan O Donoghue, Pipeline Research Limited, Glasgow
 PIG MOTION AND DYNAMICS IN COMPLEX GAS NETWORKS Dr Aidan O Donoghue, Pipeline Research Limited, Glasgow A model to examine pigging and inspection of gas networks with multiple pipelines, connections and
PIG MOTION AND DYNAMICS IN COMPLEX GAS NETWORKS Dr Aidan O Donoghue, Pipeline Research Limited, Glasgow A model to examine pigging and inspection of gas networks with multiple pipelines, connections and
Generating Power in the Pool: An Analysis of Strength Conditioning and its Effect on Athlete Performance
 Generating Power in the Pool: An Analysis of Strength Conditioning and its Effect on Athlete Performance 1 Introduction S.D. Hoffmann Carthage College shoffmann@carthage.edu November 12 th, 2014 Abstract
Generating Power in the Pool: An Analysis of Strength Conditioning and its Effect on Athlete Performance 1 Introduction S.D. Hoffmann Carthage College shoffmann@carthage.edu November 12 th, 2014 Abstract
Instruction Manual. Pipe Friction Training Panel
 Instruction Manual HL 102 Pipe Friction Training Panel 100 90 80 70 60 50 40 30 20 10 HL 102 Instruction Manual This manual must be kept by the unit. Before operating the unit: - Read this manual. - All
Instruction Manual HL 102 Pipe Friction Training Panel 100 90 80 70 60 50 40 30 20 10 HL 102 Instruction Manual This manual must be kept by the unit. Before operating the unit: - Read this manual. - All
ANNUAL GROUND TEMPERATURE MEASUREMENTS AT VARIOUS DEPTHS
 ANNUAL GROUND TEMPERATURE MEASUREMENTS AT VARIOUS DEPTHS Georgios Florides and Soteris Kalogirou Higher Technical Institute, P. O. Box 423, Nicosia 2152, Cyprus Tel. +357-22-406466, Fax: +357-22-406480,
ANNUAL GROUND TEMPERATURE MEASUREMENTS AT VARIOUS DEPTHS Georgios Florides and Soteris Kalogirou Higher Technical Institute, P. O. Box 423, Nicosia 2152, Cyprus Tel. +357-22-406466, Fax: +357-22-406480,
bespoke In general health and rehabilitation Breath-by-breath multi-functional respiratory gas analyser In human performance
 Introduction Expired Gas Analysis or indirect calorimetry, can be used to measure ventilation and the fractions of oxygen and carbon dioxide in expired air. From these measurements, the body's oxygen consumption
Introduction Expired Gas Analysis or indirect calorimetry, can be used to measure ventilation and the fractions of oxygen and carbon dioxide in expired air. From these measurements, the body's oxygen consumption
Transpiration. DataQuest OBJECTIVES MATERIALS
 Transpiration DataQuest 13 Water is transported in plants, from the roots to the leaves, following a decreasing water potential gradient. Transpiration, or loss of water from the leaves, helps to create
Transpiration DataQuest 13 Water is transported in plants, from the roots to the leaves, following a decreasing water potential gradient. Transpiration, or loss of water from the leaves, helps to create